Revealing the Key Redox Regulatory Mechanisms of ERO1A in Cancer Progression and Its Therapeutic Prospects
Published: 05 March 2025 | https://doi.org/10.63174/xdi.ZNAL8948
Abstract
Oxidative stress plays a dual role in cancer, influencing both tumor promotion and suppression through reactive oxygen species (ROS) regulation. Cancer cells upregulate antioxidant systems, evade ROS-induced cell death, and use ROS signaling to drive tumor progression and immune evasion. ERO1A (Endoplasmic Reticulum Oxidoreductase 1 Alpha), a key enzyme in redox homeostasis, is highly expressed in various cancers and has been implicated in enhancing ROS levels, thereby promoting tumor growth, metabolic reprogramming, and immune escape.This study systematically analyzed ERO1A expression across multiple cancer types using TCGA data. ERO1A was found to be significantly upregulated in cancers such as breast, liver, and lung adenocarcinoma, and its expression was correlated with poor patient survival. Functional analysis revealed that ERO1A regulates critical oxidative stress-related pathways, including lipid metabolism and the cytochrome P450 pathway, suggesting a central role in tumor cell adaptation to oxidative environments. Additionally, ERO1A's high expression was associated with increased immune-suppressive cell infiltration, supporting its role in promoting immune evasion and highlighting ERO1A as a potential diagnostic and prognostic marker in ROS-dependent cancers. By targeting the ERO1A-mediated redox balance, novel therapeutic strategies can be developed to counteract cancer progression. These results offer new insights into the interplay between oxidative stress and cancer, with ERO1A at the center of redox regulation.
Introduction
Cancer is one of the leading causes of death worldwide and is characterized by high incidence and mortality rates. Despite significant advances in treatment, the incidence of cancer continues to increase. Many cancers often present with no obvious symptoms in the early stages, resulting in late-stage diagnoses for most patients, which leads to poor treatment outcomes and low survival rates. Currently, effective early screening and diagnostic biomarkers for many types of cancer are lacking, making early detection and intervention highly challenging. Although we have gained some understanding of the etiology and mechanisms of cancer, many pathogenic mechanisms remain unclear, particularly those involving the tumor microenvironment, metabolic regulation, and immune evasion.
Redox reactions are key processes that regulate cellular homeostasis, including cell proliferation, metabolism, and signaling. In normal cells, reactive oxygen species (ROS) levels are tightly controlled and balanced by antioxidant mechanisms. However, in cancer, redox imbalance leads to ROS accumulation, promoting DNA damage, protein oxidation, and gene mutations, thereby accelerating tumor initiation and progression. Redox reactions play a central role in the onset and development of cancer. Intracellular ROS are key products of redox reactions and play important roles in maintaining cellular functions, signal transduction, and metabolic regulation[1, 2]. However, redox imbalance, particularly excessive ROS accumulation, can lead to DNA damage, protein oxidation, and gene mutations, which collectively drive cancer initiation and progression[3]. At low concentrations, ROS can act as signaling molecules for cell proliferation, activate pro-tumor signaling pathways such as PI3K/AKT and MAPK, and promote tumor growth by stimulating angiogenesis and cell migration[4]. However, high ROS levels are cytotoxic to cancer cells, limiting tumor spread by inducing apoptosis or autophagy[5]. Cancer cells adapt to high ROS conditions by activating antioxidant systems such as the Nrf2-KEAP1 pathway, which protects them from excessive ROS damage, allowing survival in a high-ROS microenvironment[6]. Redox imbalance has been recognized as one of the driving forces behind cancer metabolic reprogramming and immune evasion[7]. In terms of metabolism, ROS accumulation enables cancer cells to reconfigure their metabolic networks, thereby enhancing glycolysis and lipid metabolism to meet the demands of rapid proliferation[8]. Additionally, ROS modulate immune cell activity within the tumor microenvironment, impairing the function of immune cells, such as T cells, thus inhibiting the immune system's recognition and elimination of cancer cells, which ultimately promotes immune evasion[9]. Therefore, redox reactions exhibit dual roles in tumor biology: on one hand, they drive the malignant progression of cancer cells, while on the other hand, they provide novel opportunities for the development of antioxidant therapies and ROS-targeting treatment strategies[10].
Endoplasmic Reticulum Oxidoreductase 1 alpha (ERO1A) is a key enzyme in maintaining cellular redox balance and plays a significant role in the regulation of protein folding, particularly during the formation of disulfide bonds in nascent polypeptide chains. It produces hydrogen peroxide (H₂O₂), thereby regulating intracellular ROS levels[11]. This not only helps cells sustain essential metabolic functions but also supports the survival and proliferation of cancer cells under oxidative stress conditions[12]. ERO1A is highly expressed in various cancers, including breast cancer, hepatocellular carcinoma, and non-small-cell lung cancer. Its overexpression is closely associated with cancer malignancy, primarily by exacerbating intracellular ROS accumulation and activating pro-tumor signaling pathways such as those involving HIF-1α and VEGF, which are critical for angiogenesis and metastasis. Studies have shown that ROS enhances the anti-apoptotic ability of cancer cells, enabling them to evade immune surveillance and promote cancer adaptability and invasiveness through metabolic reprogramming[13]. Its role in the immune microenvironment is also receiving increasing attention, as high ROS levels are associated with immunosuppressive phenomena, especially by inhibiting T-cell activity, thereby promoting immune evasion in tumors. The role of ERO1A in the immune microenvironment highlights its potential as a target for cancer therapies. Targeting ERO1A or its associated redox pathways may effectively enhance the efficacy of cancer treatment by inhibiting ROS production[14].
The present study aimed to elucidate the mechanistic role of ERO1A in various cancers, particularly its function in regulating redox balance, metabolic reprogramming, and the tumor immune microenvironment. Through a comprehensive analysis of TCGA database, we explored the expression profile of ERO1A across different cancer types and assessed its value as a potential diagnostic biomarker and prognostic predictor. ERO1A not only influences oxidative stress and the survival capability of cancer cells but may also drive cancer progression by modulating immune evasion and metabolic pathways. This study provides new insights into the application of redox reactions in cancer therapy through systematic data analysis and clinical correlation studies.
Materials and Methods
Data Source and Sample Selection
This study utilized publicly available data from The Cancer Genome Atlas (TCGA), which includes RNA sequencing (RNA-seq) data from various cancer types such as breast cancer (BRCA), liver hepatocellular carcinoma (LIHC), lung adenocarcinoma (LUAD), and colon adenocarcinoma (COAD). The samples were selected based on the availability of comprehensive clinical information and RNA-seq profiles. Patients lacking complete clinical records or expression data were excluded. Only tumor and matched normal tissue samples with sufficient quality metrics were used in the analysis. 2
Gene Expression Data Analysis
The expression levels of ERO1A across different cancer types were analyzed using R software (v4.0.5). Differential expression analysis between tumor and normal tissues was conducted using the DESeq2 package, where RNA-seq data were normalized and log2 fold-change values were calculated. Statistical significance was defined as P < 0.05. For differential expression analysis, we applied Benjamini-Hochberg correction to control the false discovery rate (FDR). Adjusted P-values (FDR < 0.05) were used to determine significance unless stated otherwise.
Survival Analysis
The relationship between ERO1A expression and patient survival was assessed using Kaplan-Meier survival curves and Cox proportional hazard regression analysis. Data regarding survival times and patient outcomes were retrieved from the TCGA clinical records. Survival curves were generated using the survival and survminer R packages, and log-rank tests were used to compare the survival differences between the high and low ERO1A expression groups. Hazard ratios (HR) were calculated with a 95% confidence interval (CI), and statistical significance was set at P < 0.05.
Gene Ontology (GO) and Pathway Enrichment Analysis
To explore the biological processes and pathways associated with ERO1A, Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses were performed on co-expressed genes. The clusterProfiler R package was used for GO analysis focusing on biological processes (BP), cellular components (CC), and molecular functions (MF). KEGG pathway enrichment was conducted using the DAVID database and the results were visualized using bar plots and bubble charts. Differences were considered significant at FDR-adjusted P-values of < 0.05.
Immune Cell Infiltration Analysis
Immune cell infiltration was evaluated using CIBERSORT, which deconvoluted bulk RNA-seq data to estimate the proportions of 22 immune cell types within tumor samples. The association between ERO1A expression and immune infiltration levels was further validated using the TIMER platform, focusing on helper T cells (Th cells) and regulatory T cells (Tregs). Correlations between ERO1A expression and immune cell infiltration were computed using Spearman’s rank correlation, with significance defined as P < 0.05.
Statistical Analysis
All statistical analyses were conducted using the R software (v4.0.5). Group comparisons of gene expression were performed using the Wilcoxon rank-sum test or t-test, as appropriate. Log-rank tests were used for survival analysis and correlations between variables were assessed using Spearman’s rank correlation. P < 0.05 was set as the threshold for statistical significance across < 0.05.
Results
Result 1: Differential Expression of ERO1A in Pan-Cancer and Its Redox-Related Functions
In our analysis of TCGA database, we investigated the differential expression of ERO1A across various cancer types (Figure 1A). The results showed that ERO1A was significantly upregulated in breast cancer (BRCA), colon adenocarcinoma (COAD), hepatocellular carcinoma (LIHC), and lung adenocarcinoma (LUAD), with expression levels markedly higher than those in normal tissues. However, in pancreatic adenocarcinoma (PAAD) and thymoma (THYM), the differences in ERO1A expression did not reach statistical significance. This heterogeneous expression suggests that ERO1A may influence tumor initiation and progression in specific cancers through distinct redox regulatory mechanisms. Further analysis of differential ERO1A expression between cancer and normal tissues (Figure 1B) indicated significant upregulation in breast cancer, lung adenocarcinoma, and hepatocellular carcinoma. Notably, as a critical endoplasmic reticulum oxidoreductase, ERO1A plays an essential role in the formation of disulfide bonds during protein folding and in the regulation of cellular redox homeostasis. Therefore, upregulation of ERO1A in these cancer types may lead to increased oxidative stress within the endoplasmic reticulum, thereby affecting tumor cell proliferation and survival. Moreover, high ERO1A expression was significantly associated with a poor prognosis in certain cancer patients (Figure 1C), particularly in LUAD and LUSC. This indicates that beyond its role as a redox enzyme in basic metabolic regulation within cells, overexpression of ERO1A may exacerbate oxidative stress, thereby promoting cancer cell growth and disease progression.
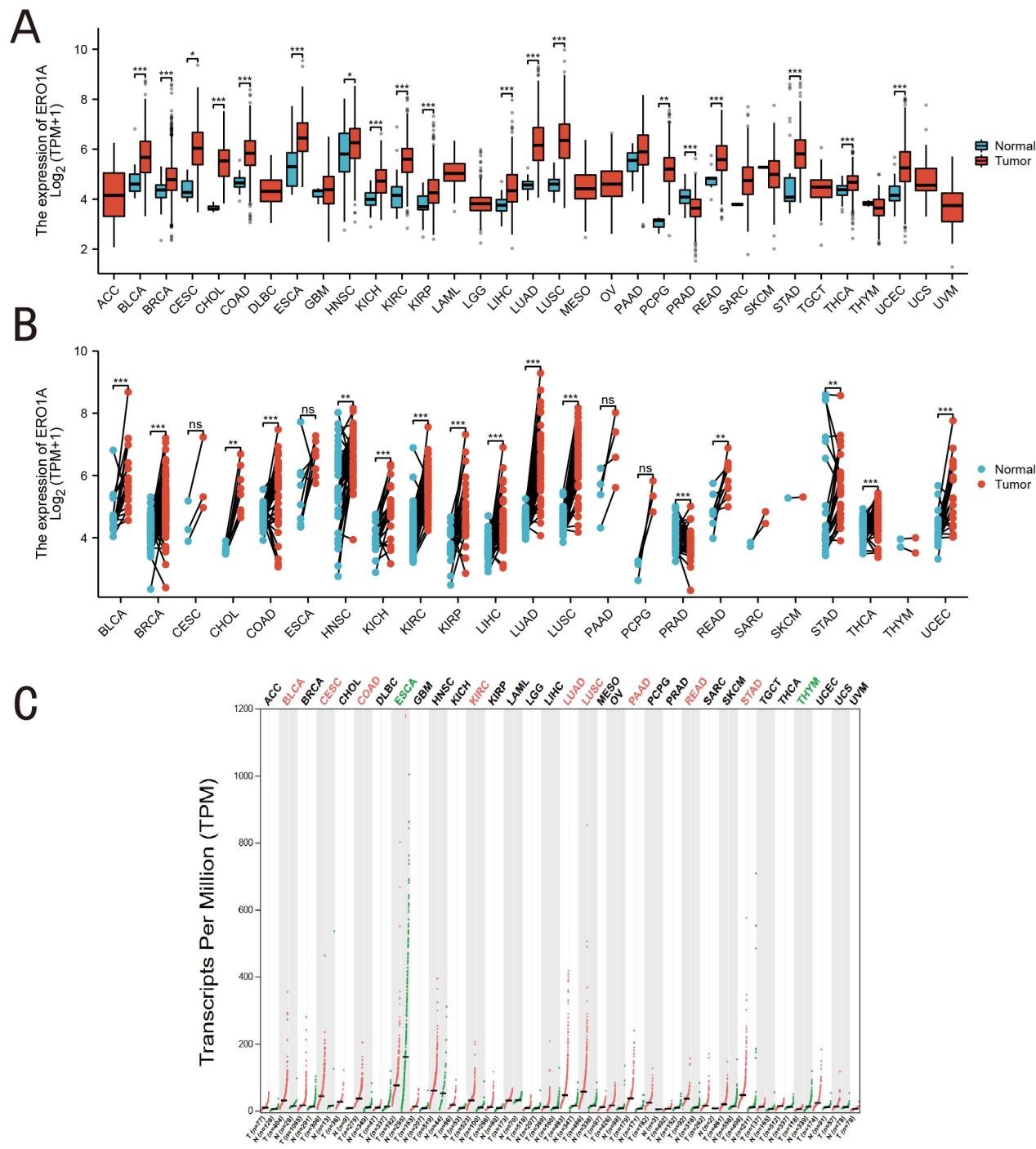
Figure 1. Expression level of ERO1A in tumors and normal tissues. A (ERO1A expression in tumor cell lines); B (ERO1A expression in TCGA tumors and adjacent normal tissues as controls); C (ERO1A expression profile across all tumor samples and paired normal tissues from GEPIA2) (*p < 0.05, **p < 0.01, ***p < 0.001).
Result 2: Association Between ERO1A Expression and Patient Survival, and the Impact of Redox Function
Kaplan-Meier survival curves and log-rank tests were used to compare overall survival between high and low ERO1A expression groups. Cox proportional hazards regression was performed with adjustments for age, gender, tumor stage, and other relevant clinical variables. The adjusted hazard ratios (HRs) showed that high ERO1A expression remained significantly associated with poor prognosis across multiple cancer types, particularly LUAD (HR = 1.87, 95% CI: 1.45–2.42, P < 0.001). These results indicate that ERO1A is an independent prognostic factor. (Figure 2A-G). This finding suggests a significant correlation between high ERO1A expression and poor prognosis. As an endoplasmic reticulum oxidoreductase, the upregulation of ERO1A may increase intracellular oxidative stress, enhancing the ability of cancer cells to cope with oxidative stress, thereby promoting malignant proliferation and anti-apoptotic characteristics. Additionally, we analyzed the effect of ERO1A expression on survival using the Cox proportional hazards regression model (Figure 2H). The analysis revealed that ERO1A was associated with a higher risk of death across several cancer types, particularly LUAD, where the hazard ratios (HR) were significantly elevated (P < 0.05). This indicates that ERO1A is not only a key regulator of redox balance, but its overexpression may also enhance cellular tolerance to endoplasmic reticulum stress within the tumor microenvironment, leading to a more aggressive cancer cell phenotype, which ultimately affects long-term patient survival. These findings collectively suggest that ERO1A, as a critical enzyme regulating redox reactions, may drive tumor progression and deterioration across multiple cancer types through the regulation of intracellular redox states. This not only supports its potential as a prognostic biomarker, but also reveals its possible role as a target in future anti-cancer therapies.
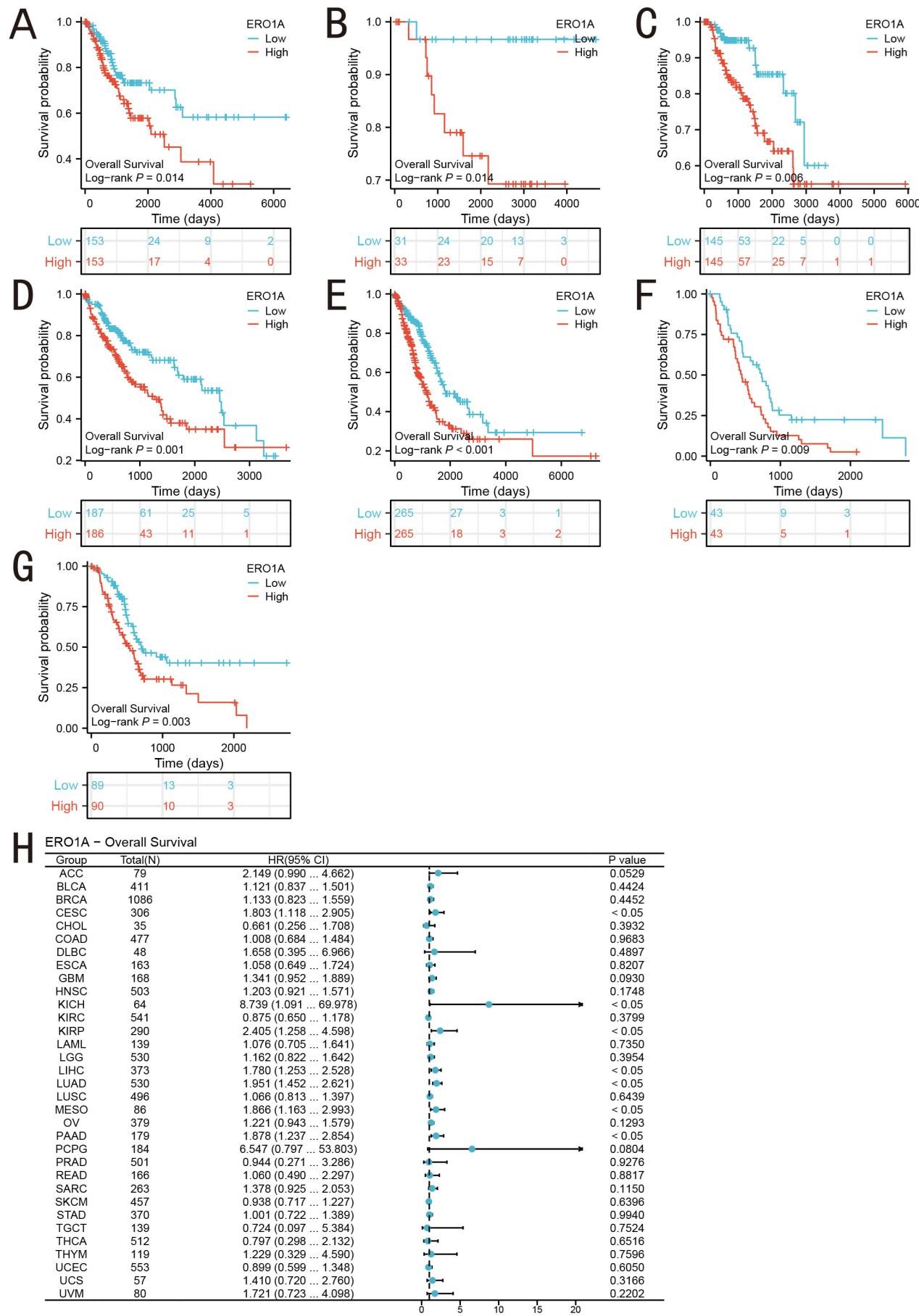
Figure 2. Prognostic Analysis of ERO1A in Different Cancer Types. (A-G) Correlation between ERO1A expression and OS prognosis A (CESC), B (KICH), C (KIRP), D (LIHC), E (LUAD), F (MESO), G (PAAD), and (H) Forest plots showing ERO1A prognostic features.
Result 3: Diagnostic Performance of ERO1A in Different Cancers and the Impact of Its Redox Function
To evaluate the potential of ERO1A as a diagnostic biomarker for cancer, we conducted a receiver operating characteristic (ROC) curve analysis and calculated the area under the curve (AUC) for different cancer types (Figure 3). The results showed that ERO1A exhibited extremely high diagnostic efficiency in cervical cholangiocarcinoma (CHOL, AUC = 0.994), Pheochromocytoma and Paraganglioma (PCPG, AUC = 0.995), and non-small cell lung cancer (NSCLC, AUC = 0.962). This suggests that ERO1A may serve as a reliable biomarker for early diagnosis of these cancers, particularly in those associated with oxidative stress, where changes in its expression levels could indicate imbalances in tumor cell metabolism and redox states. We further performed a time-dependent ROC curve analysis to assess the prognostic predictive ability of ERO1A in different cancer types (Figure 4). The results demonstrated that high ERO1A expression was significantly associated with poor prognosis in Kidney Chromophobe (KICH), cervical cancer (CESC), and pancreatic adenocarcinoma (PAAD), with high AUC values at the 1-year and 3-year survival points. These data suggest that ERO1A not only serves as a diagnostic tool but also effectively predicts patient survival, particularly in cancers associated with redox-related metabolic dysregulation. Abnormal ERO1A expression may exacerbate redox imbalances in cancer cells and influence disease progression. As an endoplasmic reticulum oxidoreductase, ERO1A's ability to regulate protein folding and redox states underscores its critical role in the tumor microenvironment. Its overexpression can enhance cellular oxidative stress responses and exacerbate oxidative damage, leading to malignant tumor progression. With its high AUC values across multiple cancers, ERO1A possesses significant diagnostic and prognostic value and could potentially serve as a target for future therapeutic strategies aimed at regulating redox reactions in tumor cells.
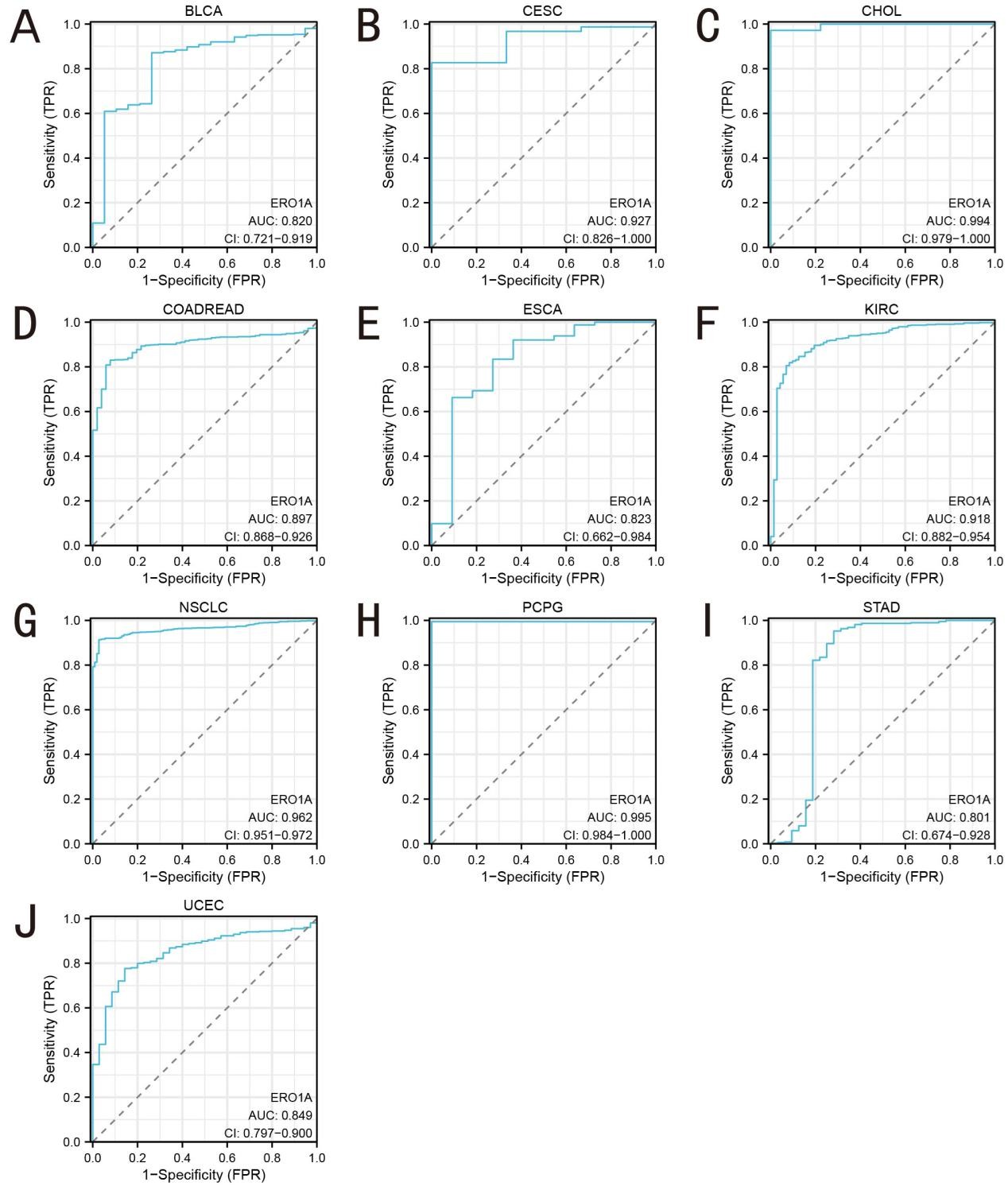
Figure 3. The diagnostic value by ROC curve of ERO1A in pan-cancer. A (BLCA); B (CESC); C (CHOL); D (COADREAD); E (ESCA); F (KIRC); G (NSCLC); H (PCPG); I (STAD), J (UCEC).
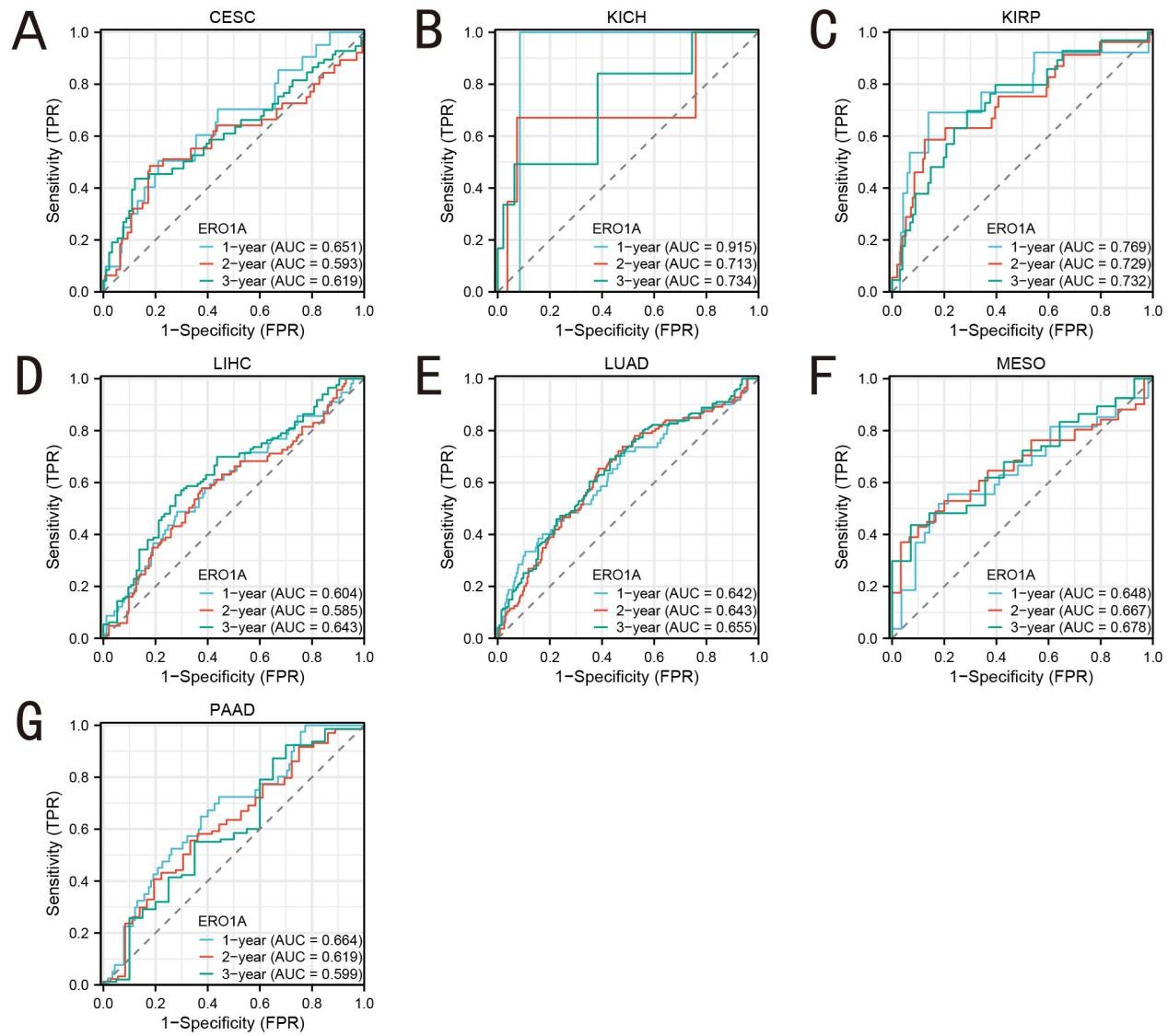
Figure 4. Area under the time-dependent ROC curve of pan-cancer. A (CESC); B (KICH); C (KIRP); D (LIHC); E (LUAD); F (MESO); G (PAAD).
Result 4: High/low expression groups in LIHC and GO/KEGG analysis
In our analysis, we identified 3,011 genes that met the screening criteria of |log2(FC)| > 1 and adjusted p-value < 0.05. Figure 5 shows the expression patterns in LIHC, along with the results from GO and KEGG pathway enrichment analyses. Under these criteria, 2,128 genes were found to be highly expressed (positive logFC), whereas 883 genes exhibited low expression (negative logFC). According to the results displayed in Figure 5B, the primary biological processes enriched by these genes included fatty acid metabolism, cell adhesion, and endoplasmic reticulum stress response. KEGG analysis showed enrichment in pathways such as drug metabolism, cytochrome P450, and fatty acid degradation, suggesting their potential involvement in metabolic reprogramming and redox balance regulated by ERO1A (Figure 5B). KEGG pathway analysis further revealed a significant enrichment of ERO1A-regulated genes in metabolic pathways, including the drug metabolism-cytochrome P450 pathway (Figure 5C). In particular, genes in the cytochrome P450 enzyme family, such as CYP2A6, CYP1A2, and CYP2B6, were significantly downregulated in cancer samples with high ERO1A expression. These enzymes are crucial for drug metabolism and detoxification, suggesting that ERO1A may influence tumor cell adaptation by modulating these pathways, potentially contributing to drug resistance. These graphical representations summarize and highlight the key biological processes and pathways enriched by the differentially expressed genes, making it easier to interpret their functional implications.
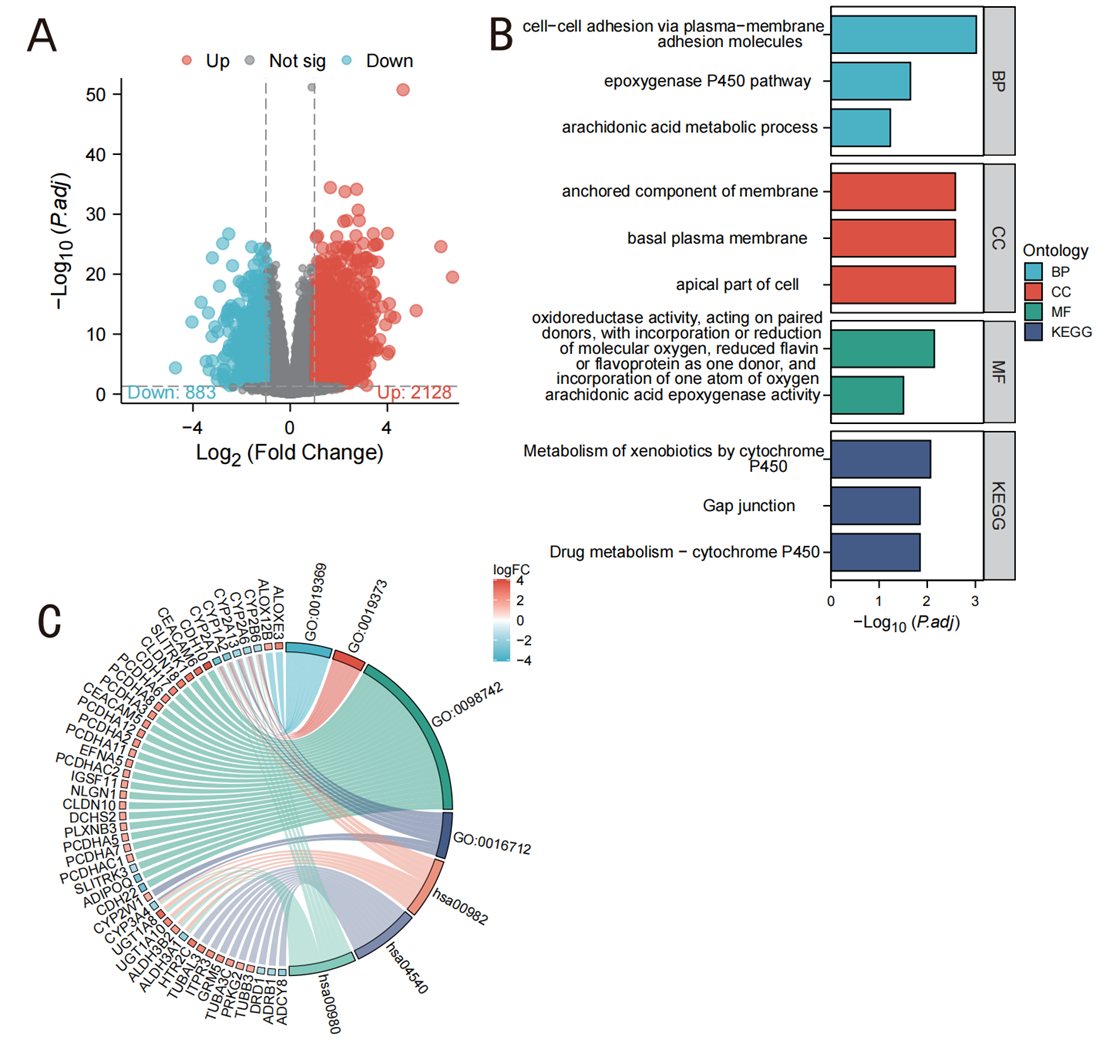
Figure 5. Expression groups in LIHC and GO/KEGG analysis. A ( Volcano map of DEGs (red: upregulation; blue: downregulation)); B (GO and KEGG enrichment analysis); C (GO enrichment analysis data distribution characteristics).
Result 5: ERO1A Co-expression and Redox Regulation in LIHC
To further investigate the molecular mechanisms of ERO1A in liver hepatocellular carcinoma (LIHC) and its impact on redox balance, we performed co-expression gene analysis (Figures 6A, 7A). Spearman correlation analysis revealed that high ERO1A expression was significantly positively correlated with multiple genes involved in redox reactions, protein folding, and cell signaling. Notably, ADAM10, EXOC5, FBXO34, and HIF1A were co-upregulated with ERO1A in LIHC. Among these, HIF1A, a critical factor in hypoxia response, showed a particularly strong correlation (R = 0.634, P < 0.001), suggesting that ERO1A may enhance cancer cell tolerance to oxidative stress by regulating hypoxia and redox homeostasis, thus promoting cancer cell survival and growth under malignant conditions (Figure 6B). Conversely, ERO1A showed a significant negative correlation with several metabolism-related genes (Figure 7B) including ACSM2A (involved in fatty acid metabolism), ADI1 (energy metabolism), and HAAO (oxidative metabolism). The downregulation of these genes in LIHC with high ERO1A expression suggests that ERO1A may drive metabolic reprogramming and redox imbalance, thereby enhancing cancer cell survival. Specifically, the downregulation of HAAO could impair antioxidant defenses, allowing cancer cells to better adapt to oxidative stress within the tumor microenvironment. As a key oxidoreductase in the endoplasmic reticulum, ERO1A regulates the formation of disulfide bonds during protein folding and plays a pivotal role in maintaining intracellular redox balance. Through co-expression gene analysis in LIHC, we demonstrated that ERO1A modulates oxidative stress and metabolic pathways, thereby contributing to cancer progression. These findings further support the potential of ERO1A as a therapeutic target for LIHC.
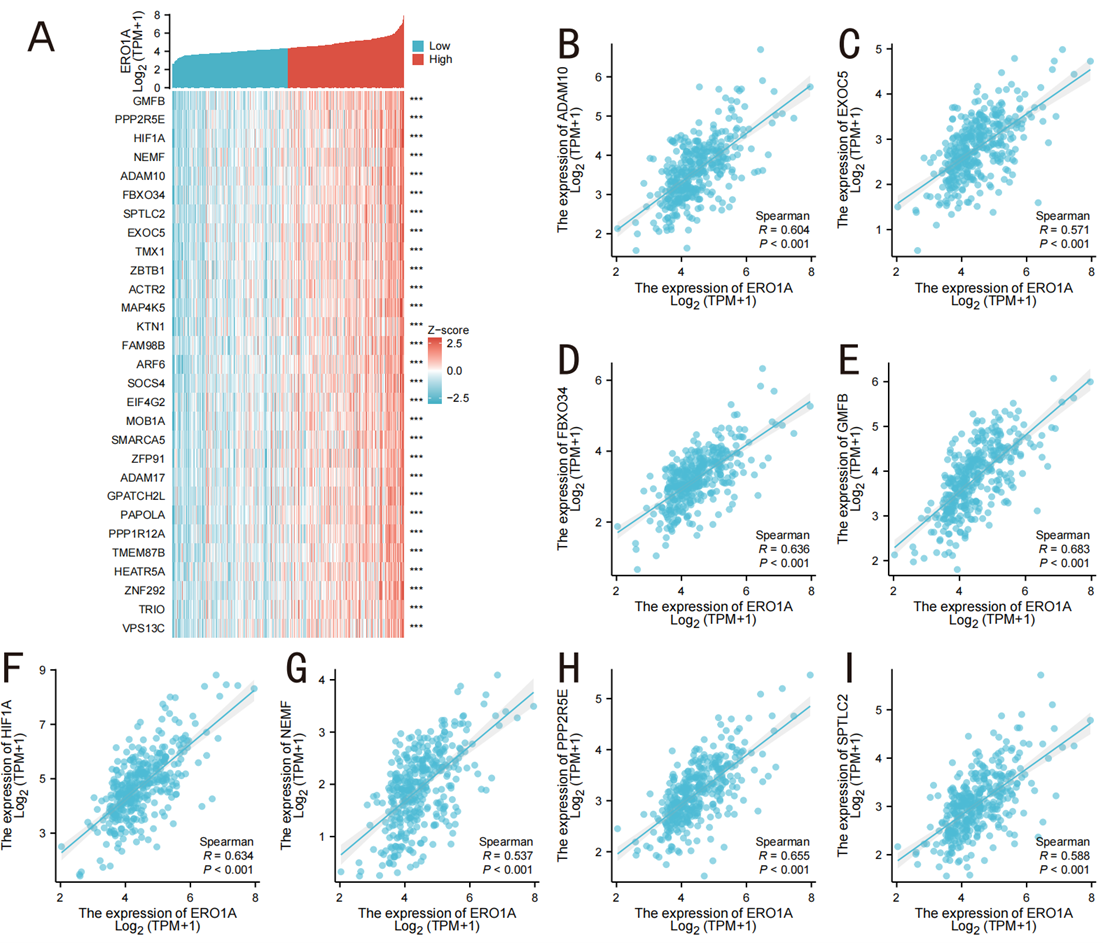
Figure 6. Top 30 genes positively correlated with ERO1A expression in LIHC. A (The gene co-expression heatmap of the top 30 genes positively correlated with ERO1A in LUAD); B-I (association analysis of the top 8 genes and ERO1A in the heatmap).
Result 6: Correlation Between ERO1A and Immune Cell Infiltration and Its Regulatory Role in Redox Reactions
To investigate the potential role of ERO1A in the tumor microenvironment, we analyzed the correlation between ERO1A expression levels and infiltration of various immune cells (Figures 8A, 8B, 8C). High ERO1A expression was significantly associated with the infiltration of helper T (Th) and Th2 cells (R = 0.299, R = 0.288, P < 0.001). Th2 cells are known to facilitate tumor immune evasion by promoting an immunosuppressive microenvironment. This finding suggests that ERO1A may indirectly suppress cytotoxic T-cell responses, reducing the efficacy of anti-tumor immunity. Future studies could investigate whether targeting ERO1A alters Th2-mediated immune suppression, potentially improving the response to immune checkpoint inhibitors.. In contrast, ERO1A also exhibited a significant negative correlation with certain immunosuppressive cells such as Th17 cells and plasmacytoid dendritic cells (pDC) (R = -0.304, R = -0.256, P < 0.001). This negative correlation might indicate that ERO1A enhances tumor invasiveness and immune evasion capabilities in a tumor microenvironment with high oxidative stress by suppressing the activity of some immunosuppressive cells. High ERO1A expression was associated with increased infiltration of Th2 cells, which are known to promote immune evasion by creating an immunosuppressive tumor microenvironment. This suggests that ERO1A may indirectly suppress cytotoxic immune responses, contributing to tumor progression. Targeting ERO1A could potentially enhance the efficacy of immunotherapy by reducing oxidative stress-induced immune suppression. Further studies are required to elucidate the precise mechanisms by which ERO1A affects immune cell activity.These findings provide new insights into the potential of developing redox-based immunotherapy targets, suggesting that ERO1A might be a key modulator of the interplay between oxidative stress and immune regulation within the tumor microenvironment.
Discussion
This study revealed significant overexpression of ERO1A in various cancers and its multiple effects on tumor development. First, through analysis of TCGA database, we found that ERO1A is significantly upregulated in several cancer types, including breast cancer, hepatocellular carcinoma, and lung adenocarcinoma. This high expression is closely associated with poor prognosis in patients with cancer, particularly among those with lower survival rates, where high ERO1A expression is significantly correlated with an increased risk of mortality. Furthermore, the high area under the curve (AUC) values for ERO1A across different cancer types indicated its high diagnostic efficacy, especially in cervical squamous cell carcinoma, kidney renal clear cell carcinoma, and non-small cell lung cancer, suggesting ERO1A's potential as a diagnostic biomarker for cancer. Based on these findings, ERO1A is not only significant for cancer diagnosis and prognostic prediction, but also plays a critical role as an endoplasmic reticulum oxidoreductase in regulating tumor redox reactions, metabolic pathways, and the tumor immune microenvironment. In summary, ERO1A influences cancer cell proliferation, metabolic reprogramming, and immune evasion by regulating the intracellular redox balance. This study provides new insights into the molecular mechanisms of ERO1A in cancer, and its potential application as a therapeutic target.
Redox reactions play a dual role in cancer initiation and progression. While moderate oxidative stress can induce apoptosis, redox imbalance in many cancer types often promotes cancer cell growth and dissemination[15]. Reactive oxygen species (ROS), as key molecules in redox reactions, when accumulated excessively, lead to DNA damage, gene mutations, increased cell proliferation, and formation of a tumor-supportive microenvironment[10]. Meanwhile, tumor cells adapt to high ROS levels by activating antioxidant systems, such as upregulating the Nrf2-KEAP1 pathway, thereby enhancing their tolerance to oxidative stress and promoting malignant transformation[6]. Oxidative stress contributes to tumor development through various mechanisms in different cancers. For example, in breast cancer, excessive ROS not only increases the rate of genetic mutations but also promotes cancer cell proliferation and angiogenesis by activating the MAPK and PI3K/AKT signaling pathways[16, 17]. Similarly, in hepatocellular carcinoma, oxidative stress-induced NF-κB activation enhances cancer cell survival, promotes inflammatory responses, and accelerates tumor dissemination by upregulating angiogenic factors[18]. These mechanisms highlight the crucial role of redox imbalances in multiple cancer types. As a key oxidoreductase in the endoplasmic reticulum, ERO1A regulates the intracellular redox status by promoting disulfide bond formation during protein folding. In this study, we found that ERO1A was significantly upregulated in breast cancer, hepatocellular carcinoma, and lung adenocarcinoma, suggesting that its overexpression may enhance ROS production, thereby promoting cancer cell proliferation and survival. This finding is consistent with recent studies indicating that excessive ERO1A may further increase ROS levels, thereby activating pro-tumor signaling pathways, such as HIF-1α and VEGF, which promote angiogenesis and tumor metastasis[19]. In other types of cancers, redox imbalance is linked not only to cancer cell proliferation and survival but also to treatment resistance. For instance, high ROS levels in pancreatic cancer cells have been shown to enhance chemotherapy resistance by upregulating the anti-apoptotic protein Bcl-2[20]. These examples demonstrate that the protumorigenic roles of ROS vary across different cancer types and are complex and multifaceted. Based on our findings, redox abnormalities promote tumor progression by activating ROS-dependent pro-tumorigenic signaling pathways, driving the metabolic reprogramming of cancer cells, and enhancing their adaptability to a malignant microenvironment. As a central factor in regulating the redox balance, ERO1A may play a crucial role in promoting malignant transformation by aiding cancer cell survival under oxidative stress conditions. Therefore, ERO1A and its regulated redox pathways are promising targets for future cancer therapy.
ERO1A regulates multiple metabolic pathways through its redox functions in the endoplasmic reticulum and plays an important role in fatty acid metabolism, the cytochrome P450 pathway, and oxidative protein folding[21, 22]. Our study further revealed that high ERO1A expression was significantly associated with the upregulation of genes involved in fatty acid metabolism, suggesting that ERO1A may facilitate cancer cell adaptation to a highly stressed microenvironment by modulating metabolic pathways. This metabolic reprogramming not only promotes cancer cell proliferation but also enhances their ability to withstand oxidative stress in a malignant microenvironment. In terms of redox regulation, ERO1A is closely involved in the formation of disulfide bonds in proteins, and its activity is mediated by protein disulfide isomerase (PDI)[23]. The ERO1A-PDI complex generates hydrogen peroxide (H₂O₂) as a by-product, thereby exacerbating intracellular oxidative stress[24]. Although moderate levels of ROS can act as signaling molecules to regulate cell proliferation and survival, excessive accumulation of ROS may trigger endoplasmic reticulum stress and apoptosis. Gene function enrichment analysis indicated that cancer cells with high ERO1A expression showed significant upregulation of redox-related genes, suggesting that disruptions in fatty acid metabolism and redox balance may enhance ROS generation and improve cancer cell tolerance to oxidative stress. This implies that ERO1A in tumors can regulate metabolic pathways via redox reactions, thereby promoting tumor progression. Moreover, the overexpression of ERO1A may also be associated with the regulation of the cytochrome P450 pathway, which could further influence drug metabolism and the survival capability of cancer cells. This mechanism suggests that targeting ERO1A and the associated redox pathways may aid in the development of new therapeutic strategies, particularly for cancers that rely on oxidative stress.
In the tumor microenvironment, redox reactions are crucial not only for the survival and proliferation of cancer cells but also for profoundly influencing tumor immune evasion through regulation of immune cell activity[3, 25]. As a key factor in regulating redox balance, high expression of ERO1A may influence the tumor immune microenvironment by enhancing ROS levels. Our study found that "high ERO1A expression was significantly positively correlated with the infiltration of various immune cells, particularly helper T (Th) and Th2 cells, suggesting that ERO1A may promote tumor immune evasion by enhancing the activity of immunosuppressive cells." This finding is consistent with existing studies, which indicate that redox imbalance not only promotes cancer cell survival, but also alters the tumor microenvironment, making it more immunosuppressive[26]. In various types of cancers, ROS modulate immune cell infiltration and function through redox regulatory networks. For instance, high ROS levels in breast cancer can promote immune evasion by inhibiting T-cell function[9]. Recent research has shown that ROS impair the anti-tumor ability of T cells by inhibiting their metabolism and function, including suppressing glycolysis, thereby weakening their proliferative and effector capabilities[27]. In our study, high ERO1A expression was associated with a reduction in immunosuppressive cells such as Th17 cells, suggesting that ERO1A may facilitate malignant progression by regulating immune evasion mechanisms. This finding further supports the potential of targeting ERO1A as a modulator of immune response. Additionally, the role of ERO1A in cancer diagnosis and prognosis is widely recognized. Kaplan-Meier survival curves and Cox regression analysis indicated that high ERO1A expression was closely associated with poor prognosis in patients with breast cancer and hepatocellular carcinoma. This aligns with the mechanisms observed in other studies, where ROS promote cancer cell resistance to apoptosis and metabolic reprogramming[28]. The high AUC values of ERO1A further support its potential as a diagnostic biomarker, particularly in cancers with elevated ROS levels, where changes in ERO1A expression could serve as an important reference for the early detection of tumor initiation and progression[13]. In PAAD and THYM, the differences in ERO1A expression did not reach statistical significance, potentially due to limited sample sizes or unique tumor microenvironment characteristics in these cancer types. For instance, pancreatic adenocarcinoma is known for its dense stromal components, which may obscure the effects of ERO1A. Similarly, the thymic environment's specialized immune functions could influence redox dynamics. These results highlight the need for cancer-specific investigations to elucidate ERO1A's role. In conclusion, ERO1A influences cancer cell survival, immune evasion, and treatment resistance by regulating the tumor immune microenvironment and redox balance. Thus, ERO1A is an important subject of study for cancer diagnosis and prognosis prediction, and as a potential therapeutic target.
This study revealed the multiple roles of ERO1A in cancers, particularly its functions in regulating redox balance, metabolic reprogramming, and the tumor immune microenvironment. However, this research primarily relies on bioinformatics analyses and lacks experimental validation. Specific mechanisms, such as how ERO1A affects immune cell function and ROS production across different cancer types, require further confirmation using in vitro and in vivo experiments. Future studies should explore the differential roles of ERO1A in various cancers, particularly ROS-dependent cancers. In particular, experimental validation could involve inhibiting ERO1A activity or upregulating antioxidant pathways to assess their direct effects on tumor growth and immune evasion. Additionally, the feasibility of targeting ERO1A as a therapeutic target should be evaluated using both in vitro and in vivo models, and its potential as a combination treatment strategy should be investigated. In the future, antioxidant therapies targeting ERO1A could be combined with other anti-cancer strategies, such as immune checkpoint inhibitors and chemotherapy, to potentially provide new therapeutic options for specific cancer patients. Potential therapeutic approaches targeting ERO1A include small-molecule inhibitors to block its enzymatic activity, RNA-based therapies like siRNA to suppress its expression, and monoclonal antibodies to disrupt its function. Challenges such as achieving specificity, effective drug delivery, and minimizing off-target effects must be addressed in future research. Combining ERO1A inhibitors with existing treatments, such as immune checkpoint inhibitors, could also be explored as a synergistic strategy. Further more future studies should involve experimental validation to confirm the findings of this study. Specific experiments could include ERO1A knockdown or overexpression in vitro to examine its effects on ROS levels and cancer cell survival, as well as in vivo models to investigate its role in tumor progression and immune evasion. This study relies on TCGA datasets, which, while comprehensive, may have inherent limitations such as sample bias and lack of uniformity in data collection. Additionally, the findings are based on bioinformatics analyses without experimental validation. Future research should focus on multi-center validation using clinical samples to confirm the diagnostic and prognostic value of ERO1A. In-depth functional studies, including animal models and patient-derived organoids, are also essential to understand the precise role of ERO1A and to explore its potential as a therapeutic target.
This study provides an in-depth analysis of ERO1A expression and its regulatory mechanisms in various cancers, revealing its significant role as an endoplasmic reticulum oxidoreductase. ERO1A promotes cancer cell survival, proliferation, and immune evasion by regulating intracellular redox balance and metabolic pathways. Particularly, in cancers that depend on high ROS levels, high ERO1A expression is closely associated with poor prognosis, highlighting its importance in cancer diagnosis, prognostic prediction, and as a potential therapeutic target. Further elucidation of the specific mechanisms of ERO1A, along with clinical validation of its feasibility as a biomarker and therapeutic target, holds promise for offering new directions and strategies for cancer diagnosis and treatment.
Conclusion
This study reveals the crucial role of ERO1A in various cancers, finding it significantly upregulated in breast, liver, and lung adenocarcinoma, with high expression linked to poor patient outcomes. Mechanistic insights show that ERO1A influences tumor progression by modulating oxidative stress and redox balance, affecting key pathways like lipid metabolism and cytochrome P450, and promoting tumor immune escape through immune cell infiltration in the tumor microenvironment. These findings suggest ERO1A as a potential biomarker for cancer prognosis and offer new targets for redox-based cancer therapies, advancing personalized treatment.
References
- Lennicke C., Cochemé H. M. "Redox metabolism: ROS as specific molecular regulators of cell signaling and function." Mol. Cell. 2021, 81, 18, 3691-3707.
- Zoccarato A., Schlattner U., Calzia D., Fornasaro S., Serpe L., Leanza G., Bisoffi M., Panfoli I. "The nexus between redox state and intermediary metabolism." FEBS J. 2022, 289, 18, 5440-5462.
- Liu W., Zhao Y., Wang X., Wang Q., Yang Y., Wang J., Zhu L., Yang Y. "Redox Dysregulation in the Tumor Microenvironment Contributes to Cancer Metastasis." Antioxid. Redox Signal. 2023, 39, 7-9, 472-490.
- Jin S. Y., Lee H. S., Kim E. K., Ha J. M., Kim Y. W., Bae S. S. "Reactive oxygen species and PI3K/Akt signaling in cancer." Free Radic. Biol. Med. 2014, 75, Suppl 1, S34-5.
- Hung S. Y., Huang W. J., Lin H. Y., Chen W. C., Wang C. C., Hour M. J., Ko B. S., Huang L. J., Tsay H. S., Hung M. C., Liu Y. M., Yuan S. F., Yang S. C., Lu C. T., Way T. D. "Rhopaloic acid A induces apoptosis, autophagy and MAPK activation through ROS-mediated signaling in bladder cancer." Phytomedicine 2021, 92, 153720.
- Panieri E., Saso L. "Inhibition of the NRF2/KEAP1 Axis: A Promising Therapeutic Strategy to Alter Redox Balance of Cancer Cells." Antioxid. Redox Signal. 2021, 34, 18, 1428-1483.
- Guo C., Shen F., Cheng Y., Wu X., Liu H., Wang H., Hu Y. "Immunometabolism: A new target for improving cancer immunotherapy." Adv. Cancer Res. 2019, 143, 195-253.
- Wang K., Fan H., Chen Q., Ma G., Zhu M., Zhang X., Zhang Y., Yu J. "Targeting Metabolic-Redox Circuits for Cancer Therapy." Trends Biochem. Sci. 2019, 44, 5, 401-414.
- Zhang W., Han L., Shi L., Shao L., Wang Y., Wang X., Liu Y., Wang Z., Qiao Y., Chen Y., Zheng Y., Yuan S., Wang C., Ma J., Cao F., Fan Z., Song X. "Multifunctional Redox-Responsive Nanoplatform with Dual Activation of Macrophages and T Cells for Antitumor Immunotherapy." ACS Nano 2023, 17, 15, 14424-14441.
- Wang Y., Qi H., Liu Y., Duan C., Liu X., Xia T., Chen D., Piao H. L., Liu H. Y. "The double-edged roles of ROS in cancer prevention and therapy." Theranostics 2021, 11, 10, 4839-4857.
- Ramming T., Okumura M., Kanemura S., Baday S., Birk J., Moes S., Spiess M., Jenö P., Bernèche S., Inaba K., Appenzeller-Herzog C. "A PDI-catalyzed thiol-disulfide switch regulates the production of hydrogen peroxide by human Ero1." Free Radic. Biol. Med. 2015, 83, 361-372.
- Chun K. S., Jang J. H., Kim D. H. "Role of Reductive versus Oxidative Stress in Tumor Progression and Anticancer Drug Resistance." Cells 2021, 10, 4, 758.
- Varone E., Decio A., Chernorudskiy A., Minoli L., Brunelli L., Ioli F., Piotti A., Pastorelli R., Fratelli M., Gobbi M., Cecconi F., Giavazzi R., Zito E. "The ER stress response mediator ERO1 triggers cancer metastasis by favoring the angiogenic switch in hypoxic conditions." Oncogene 2021, 40, 9, 1721-1736.
- Zhang J., Lei Z., Huang Z., Zhang X., Zhou Y., Luo Z., Zeng W., Su J., Peng C., Chen X. "Small molecules regulating reactive oxygen species homeostasis for cancer therapy." Med. Res. Rev. 2021, 41, 1, 342-394.
- Kaneko M., Sasayama T., Tanaka K., Yamazaki Y. "Cell-Membrane Permeable Redox Phospholipid Polymers Induce Apoptosis in MDA-MB-231 Human Breast Cancer Cells." Biomacromolecules 2019, 20, 12, 4447-4456.
- Huang J. S., Cho C. Y., Hong C. C., Liu Y. C., Hsu F. C., Cheng Y. W., Chou M. T., Yang Y. H., Lin W. Y. "Oxidative stress enhances Axl-mediated cell migration through an Akt1/Rac1-dependent mechanism." Free Radic. Biol. Med. 2013, 65, 1246-1256.
- Tsai C. H., Shen Y. C., Chen H. W., Liu K. L., Chang J. W., Chen P. Y., Lin S. Y., Mok K. T., Yang L. X., Wang J. Y., Chang Y. H., Li C. Y., Chien C. C., Huang S. F. "Docosahexaenoic acid increases the expression of oxidative stress-induced growth inhibitor 1 through the PI3K/Akt/Nrf2 signaling pathway in breast cancer cells." Food Chem. Toxicol. 2017, 108, Part A, 276-288.
- Payne C. M., Weber C., Crowley-Skillicorn C., Dvorak K., Bernstein H., Bernstein C., Holubec H., Dvorakova B., Garewal H. "Deoxycholate induces mitochondrial oxidative stress and activates NF-kappaB through multiple mechanisms in HCT-116 colon epithelial cells." Carcinogenesis 2007, 28, 1, 215-222.
- Wang N., Zhan T., Ke T., Huang X., Ke D., Wang Q., Li H. "Increased expression of RRM2 by human papillomavirus E7 oncoprotein promotes angiogenesis in cervical cancer." Br. J. Cancer 2014, 110, 4, 1034-1044.
- Zhou J., Du Y. "Acquisition of resistance of pancreatic cancer cells to 2-methoxyestradiol is associated with the upregulation of manganese superoxide dismutase." Mol. Cancer Res. 2012, 10, 6, 768-777.
- Jha V., Jha N. N., Ghosh K. S., Lenz S. M., Cole M. P., Townsend D. M. "ERO1-PDI Redox Signaling in Health and Disease." Antioxid. Redox Signal. 2021, 35, 13, 1093-1115.
- Uchida Y., Nishi K., Fujisawa T., Kawashima Y., Manabe S., Saito Y., Hashimoto M., Enomoto M., Mikasa M., Honda A., Ikeda K., Arai H., Lee K. F., Saito K., Kanno K., Shimano H., Nakagawa Y. "Trans-2-enoyl-CoA reductase limits Ca2+ accumulation in the endoplasmic reticulum by inhibiting the Ca2+ pump SERCA2b." J. Biol. Chem. 2021, 296, 100310.
- Matsusaki M., Okuda A., Masuda T., Koishihara K., Mita R., Iwasaki K., Hara K., Naruo Y., Hirose A., Tsuchiya Y., Mizobata T., Kawata Y. "Functional Interplay between P5 and PDI/ERp72 to Drive Protein Folding." Biology 2021, 10, 11, 1112.
- Cheng F., Geng L., Wang Y., Chen W., Wu J., Liu X., Liu S., Wu J., Luo C., Zhu E., Duan Y., Liu X., Liu X., Zito E. "Reducing oxidative protein folding alleviates senescence by minimizing ER-to-nucleus H2O2 release." EMBO Rep. 2023, 24, 8, e56439.
- Xing F., Jia M., Wang K., Liu J., Gao Z., Cao S., Liu X. "The Relationship of Redox With Hallmarks of Cancer: The Importance of Homeostasis and Context." Front. Oncol. 2022, 12, 862743.
- Aboelella N. S., Brandle C., Kim T., Ding Z. C., Zhou G. "Oxidative Stress in the Tumor Microenvironment and Its Relevance to Cancer Immunotherapy." Cancers 2021, 13, 5, 986.
- Daneshmandi S., Wegiel B., Seth P. "Blockade of 6-phosphogluconate dehydrogenase generates CD8+ effector T cells with enhanced anti-tumor function." Cell Rep. 2021, 34, 10, 108831.
- Liu L., Yu X., Li S., Wang P., Yu P., Wang C., Du Q., Meng Q., Yu Q., Liu J., Huang X., Tian Z., Wen Y., Wu K., Yang J., Liu Y., Ding Y., Liu X., Zhang Y., Chen D., Wu L., Yang L., Bai C., Wang J., Gu C., Lu Z. "ERO1L Is a Novel and Potential Biomarker in Lung Adenocarcinoma and Shapes the Immune-Suppressive Tumor Microenvironment." Front. Immunol. 2021, 12, 677169.
Acknowledgements
Funding Statement: No funding.
Ethical Compliance: All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki Declaration and its later amendments or comparable ethical standards.
Conflict of Interest declaration: The authors declare that they have no affiliations with or involvement in any organization or entity with any financial interest in the subject matter or materials discussed in this manuscript.