Gut Microbiota and Thyroid Diseases: A Comprehensive Review of Mechanisms and Clinical Implications
published: 18 September 2025 | https://doi.org/10.63174/xdi.BDAP6489
Abstract
The interplay between gut microbiota and thyroid diseases has emerged as a cutting-edge topic in endocrinology and microbiome research. Accumulating evidence indicates that gut microbes not only influence thyroid hormone synthesis and metabolism through metabolites such as short-chain fatty acids and bile acids, but also play critical roles in immune homeostasis, thereby contributing to the pathogenesis and progression of Graves' disease, Hashimoto's thyroiditis, thyroid dysfunction, and thyroid cancer. Epidemiological and clinical studies suggest that specific microbial signatures are associated with disease risk, subtype classification, therapeutic response, and recurrence, highlighting their potential as diagnostic and prognostic biomarkers. Emerging interventions, including probiotics, dietary modulation, and fecal microbiota transplantation, have shown promising effects in animal models and preliminary clinical studies, offering new opportunities for personalized management. Nevertheless, current research faces notable challenges: causal relationships remain unclear, disease heterogeneity and population differences compromise reproducibility, and biases inherent to low-biomass samples and sequencing methodologies limit evidence strength. Future investigations should focus on standardized protocols, cross-cohort validation, multi-omics integration, and mechanistic trials to elucidate the biological basis of the “gut–thyroid axis” and accelerate its translation into clinical diagnostics, risk stratification, and therapeutic interventions.
1. Introduction
In recent years, the relationship between gut microbiota and thyroid diseases has become a major focus of both basic and clinical research. Growing evidence indicates that the composition of gut microbiota is closely associated with the onset and progression of various thyroid disorders, and that microbial dysbiosis may promote disease pathology through disrupted metabolic pathways and abnormal immune regulation [1, 2]. On this basis, the concept of the “gut–thyroid axis” has been proposed, providing a new theoretical framework for understanding the mechanisms underlying thyroid diseases and laying the foundation for exploring potential diagnostic and therapeutic strategies. At the metabolic level, gut microbiota regulate the production of short-chain fatty acids (SCFAs), thereby contributing to immune homeostasis and thyroid hormone metabolism. Studies have shown that in patients with thyroid diseases, beneficial taxa such as Bifidobacterium and Lactobacillus are markedly reduced, whereas potential pathogenic bacteria including Bacteroides and Proteobacteria are significantly increased [1, 3]. Such dysbiosis may lead to decreased SCFA levels, which in turn impair mucosal immune function, alter the inflammatory microenvironment, and further affect the synthesis, transport, and metabolism of thyroid hormones [4]. From an immunological perspective, gut microbiota modulate the differentiation and function of host immune cell subsets, thereby contributing to the development and progression of autoimmune thyroid diseases (AITD). Some studies suggest that specific gut microbes can influence the immune response in Graves' disease by altering the proportion of CD4⁺ T cells, thereby exacerbating disease progression[3]. This effect may be indirectly mediated through the “gut–thyroid axis,” in which microbial compositional changes feedback to influence thyroid hormone synthesis and metabolism[5]. In terms of clinical interventions, modulation of gut microbiota has been considered a promising approach for the management of thyroid disorders. Probiotic supplementation has demonstrated beneficial effects in improving thyroid function and alleviating clinical symptoms. For example, strains such as Lactiplantibacillus plantarum and Bifidobacterium longum have been shown to restore microbial homeostasis and improve patient outcomes when combined with conventional therapies. In addition, fecal microbiota transplantation (FMT), as an emerging strategy, has demonstrated the potential to reshape gut microbiota in both Graves' disease animal models and preliminary clinical studies. This effect may occur via the gut–thyroid–brain axis, through the regulation of immune responses and micronutrient metabolism, thereby influencing disease progression. In summary, current evidence supports a critical role for gut microbiota in the pathogenesis and progression of thyroid diseases, and highlights their potential as targets for individualized therapy and integrated management. Nevertheless, the specific mechanistic roles of distinct microbial taxa remain insufficiently characterized, and their feasibility as diagnostic, prognostic, and therapeutic tools requires further validation [2, 6].
2. Mechanistic and Clinical Evidence Linking Gut Microbiota to Thyroid Diseases
To provide an integrative overview before discussing disease-specific mechanisms, we present a schematic illustration of the gut–thyroid axis (Figure 1). This framework highlights how gut microbiota, including both beneficial and pathogenic taxa, influence thyroid health via microbial metabolites such as short-chain fatty acids (SCFAs), lipopolysaccharides (LPS), secondary bile acids, and indoles. These intermediates modulate key mechanistic pathways, including immune regulation, nutrient metabolism, drug handling, and barrier function, ultimately contributing to distinct thyroid disease phenotypes such as Graves' disease, Hashimoto's thyroiditis, hypothyroidism, hyperthyroidism, and thyroid cancer. The figure also underscores clinical implications, ranging from diagnostic biomarkers to therapeutic interventions such as probiotics, dietary modulation, and fecal microbiota transplantation (Figure 2).
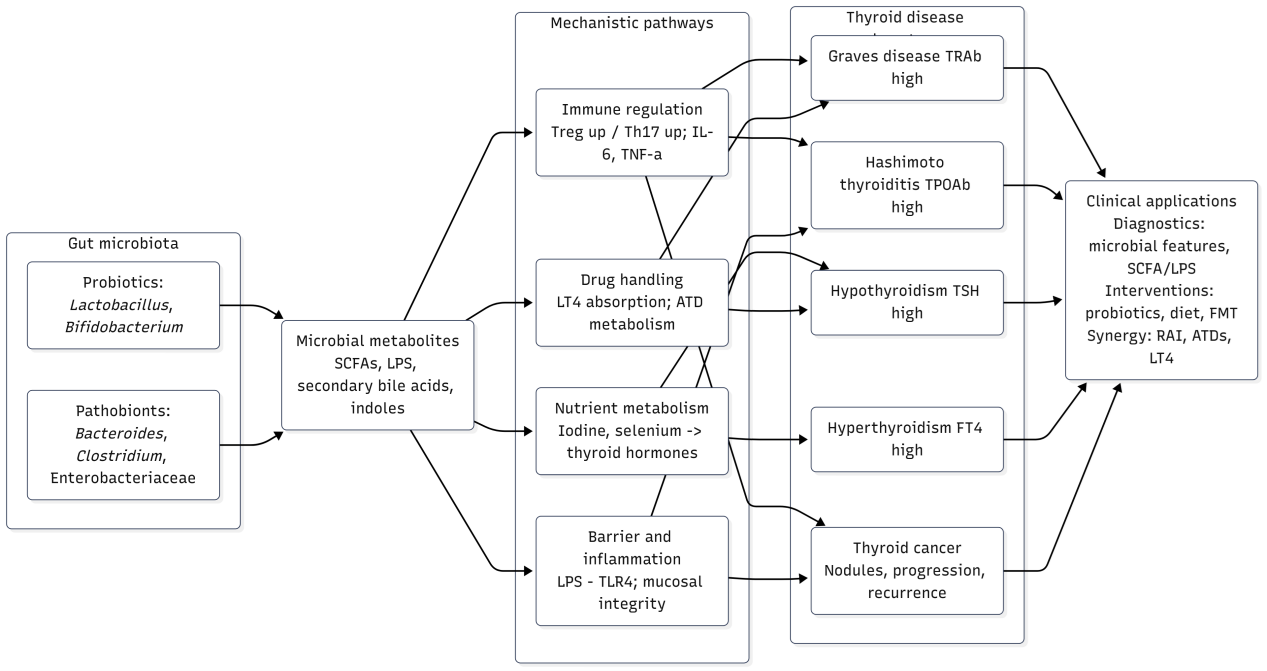
Figure 1. The gut–thyroid axis: mechanisms and clinical implications. Schematic illustration showing how gut microbiota (probiotics such as Lactobacillus and Bifidobacterium; pathobionts including Bacteroides, Clostridium, and Enterobacteriaceae) and their metabolites (short-chain fatty acids, LPS, secondary bile acids, indoles) influence thyroid diseases through multiple mechanistic pathways. These include immune regulation (Treg/Th17 balance, cytokines such as IL-6 and TNF-α), nutrient metabolism (iodine and selenium utilization), barrier integrity and inflammation (LPS–TLR4 signaling), and drug handling (levothyroxine absorption, antithyroid drug metabolism). The resulting alterations contribute to distinct thyroid disease phenotypes, including Graves’ disease (↑TRAb), Hashimoto thyroiditis (↑TPOAb), hypothyroidism (↑TSH), hyperthyroidism (↑FT4), and thyroid cancer (nodules, progression, recurrence). These mechanistic insights highlight clinical applications in diagnostics (microbial features, SCFA/LPS profiling), interventions (probiotics, diet, fecal microbiota transplantation), and therapeutic synergy with standard treatments (radioactive iodine, antithyroid drugs, levothyroxine).
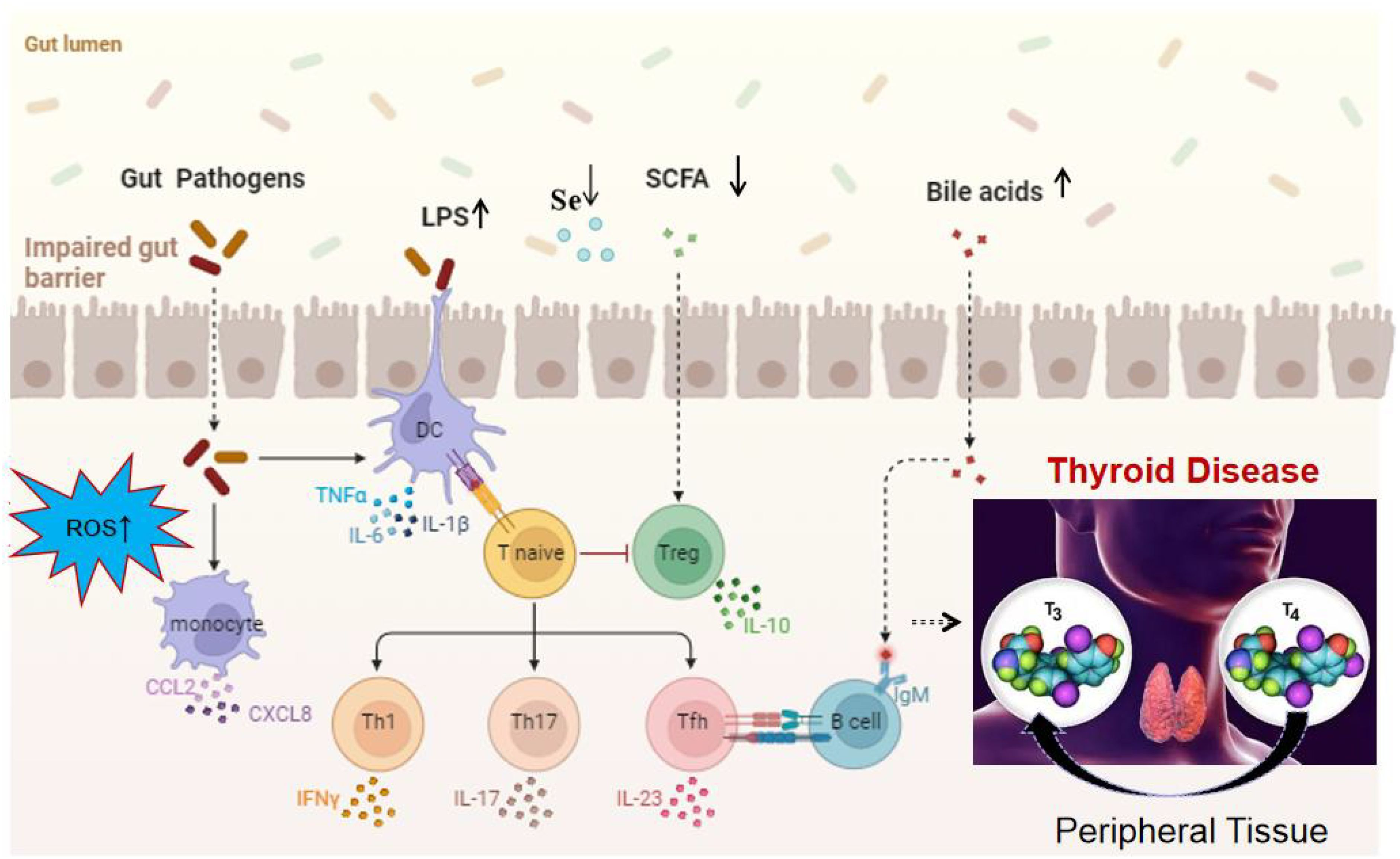
Figure 2. Mechanistic overview of the impact of gut microbiota on thyroid function[1] (licensed under CC BY 4.0.).
2.1. Gut Microbiota and Thyroid Hormone Metabolism
The relationship between gut microbiota and thyroid hormone metabolism has emerged as a research hotspot in recent years. As a complex microbial ecosystem of the human body, the gut microbiota not only participates in digestion and nutrient metabolism but also regulates endocrine homeostasis through multiple pathways. Emerging evidence suggests that gut microbes can directly or indirectly influence the synthesis, transport, and metabolism of thyroid hormones, thereby contributing to the development and progression of thyroid disorders. One important mechanism involves the interaction between bile acids and gut microbiota. The diversity of bile acids results from the combined activity of the host and intestinal microbes. Microbes regulate bile acid deconjugation, transformation, and recirculation, while bile acid composition in turn shapes microbial diversity and stability. This bidirectional interaction has been implicated in diseases such as inflammatory bowel disease, colorectal cancer, hepatocellular carcinoma, type 2 diabetes, and polycystic ovary syndrome[7]. Given that bile acids influence the activity of thyroid hormone-binding proteins and metabolic pathways, the microbiota–bile acid axis may indirectly affect thyroid hormone homeostasis by modulating gut stability and immune responses. Gut dysbiosis has also been associated with primary hypothyroidism. Compared with healthy individuals, patients with hypothyroidism exhibit significant differences in both α- and β-diversity of gut microbiota. Specific genera such as Veillonella, Paraprevotella, Neisseria, and Rheinheimera were found to distinguish untreated hypothyroidism patients from healthy controls with high accuracy[8]. These structural changes may reduce the capacity for short-chain fatty acid (SCFA) production while increasing serum lipopolysaccharide (LPS) levels, thereby inducing systemic inflammation and impairing thyroid hormone synthesis and function. Further evidence arises from studies on the interplay among nutrition, gut microbiota, and thyroid hormones. Iodine and selenium are essential micronutrients for thyroid hormone synthesis and activity, and the gut microbiota plays a critical role in their bioavailability and metabolism. Certain probiotics, such as Lactobacillus and Bifidobacterium, enhance the bioavailability of selenium metabolites and thereby indirectly support thyroid hormone synthesis. In contrast, dysbiosis may reduce nutrient absorption efficiency and exacerbate thyroid hormone imbalance[9]. Moreover, microbial metabolites themselves may modulate thyroid hormone activity. For example, SCFAs can affect the activity of deiodinases, thereby influencing the conversion of thyroxine (T4) into triiodothyronine (T3). Other metabolites such as indole derivatives and secondary bile acids may interfere with thyroid hormone metabolism indirectly by binding to nuclear receptors including FXR and PXR[10]. In summary, gut microbiota influence thyroid hormone metabolism through multiple levels of regulation, including bile acid modulation, dysbiosis-associated hypothyroidism risk, nutrient metabolism, and microbial metabolite-mediated control of hormone conversion. These findings not only provide new insights into the pathophysiology of thyroid disorders but also highlight the microbiota as a potential therapeutic target. Future studies should leverage longitudinal cohorts and multi-omics analyses to clarify the role of specific taxa and metabolic pathways in thyroid hormone regulation, thereby advancing the development of personalized intervention strategies.
2.2. Gut Microbiota and Graves' Disease: Mechanisms and Clinical Evidence
Graves' disease (GD) is one of the most common autoimmune thyroid disorders, characterized by limited treatment efficacy, high relapse rates, and frequent adverse effects. Increasing evidence suggests that gut microbiota play an important role in its pathogenesis. Clinical studies have shown that gut microbial diversity is markedly reduced in GD patients, with a significant decrease in Firmicutes and an increase in Bacteroidetes[11, 12]. These compositional shifts are closely associated with immune dysregulation, indirectly supporting the concept of the gut–thyroid axis. Beyond disease onset, gut microbiota also demonstrate diagnostic potential. High-throughput sequencing has identified microbial signatures strongly associated with GD, which can differentiate GD from other conditions (e.g., Parkinson's disease) with high specificity and predictive value [13]. Such noninvasive microbial biomarkers may enable earlier detection and individualized management. Importantly, the microbiota–GD relationship appears bidirectional. Bidirectional Mendelian randomization analyses indicate that certain taxa may act as risk factors for GD, whereas others may exert protective effects [14–16]. This dual causality underscores the complexity of the gut–thyroid axis in GD pathogenesis. Compelling experimental evidence comes from microbial intervention studies. A recent investigation in a GD mouse model evaluated the effects of the commensal bacterium Bacteroides fragilis and its metabolite propionate. Oral supplementation with either B. fragilis or propionate significantly reduced serum inflammatory cytokines, total thyroxine (TT4), and TSH receptor antibody (TRAb) levels, while decreasing circulating Th17 cells and increasing regulatory T cells (Tregs). These changes alleviated systemic inflammation, hyperthyroidism, and TSHR autoimmunity. Histological analysis further revealed reduced proportions of M1 macrophages and increased M2 macrophages in thyroid tissue, accompanied by decreased inflammation and thyroid size. Notably, the combination of B. fragilis or propionate with methimazole (MMI) produced synergistic effects, ameliorating pathological damage while reducing the required MMI dose and minimizing drug-related adverse effects. This study provided the first experimental evidence that gut microbiota and their metabolites can serve as effective adjuvant strategies for GD, offering a microbiota-based rationale for personalized interventions [17]. In summary, current research highlights multiple roles of the gut microbiota in GD, spanning dysbiosis, immune dysfunction, diagnostic potential, and therapeutic efficacy. Future studies should clarify the causal contributions of specific taxa and evaluate the clinical feasibility of microbiota- and metabolite-based interventions to establish more comprehensive and precise strategies for GD management.
2.3. Gut Microbiota and Hashimoto's Thyroiditis: Mechanisms and Clinical Evidence
The relationship between gut microbiota and Hashimoto's thyroiditis (HT) has attracted increasing attention in recent years. Evidence indicates that the gut microbiota plays a pivotal role in host immune regulation and is closely linked to the pathogenesis of multiple autoimmune diseases. HT, the most common autoimmune thyroid disorder, arises from a multifactorial etiology involving both genetic and environmental factors. Growing evidence shows that the composition and abundance of gut microbiota in HT patients differ significantly from healthy controls, suggesting that microbial alterations may influence disease onset and progression through specific mechanisms[18]. Multi-omics studies have further revealed characteristic microbial signatures in HT. Integrated metagenomic and host transcriptomic analyses demonstrated reduced microbial diversity, with early HT patients showing increased abundance of Bacillota_A and Spirochaetota, along with significant differences across 24 genera and 67 species[19]. Functional analyses indicated altered activity of immune- and infection-related pathways, providing new insights into the molecular mechanisms of HT[20]. Such changes may disrupt peripheral thyroid homeostasis, thereby promoting autoimmune responses and disease progression[21]. Moreover, interactions between the microbiota and host transcriptome were found to involve metabolic, immune, and cancer-related pathways, further reinforcing the role of the gut–thyroid axis in HT[2]. Clinical and experimental data also support this association. A case–control study reported that butyrate-producing bacteria were markedly reduced in HT patients, while opportunistic pathogens such as Escherichia/Shigella were enriched. This imbalance correlated with elevated pro-inflammatory cytokines and higher titers of anti-thyroid antibodies[22]. In an animal model, oral supplementation with Lactobacillus reuteri significantly lowered thyroiditis scores and anti-thyroid peroxidase antibody (TPOAb) levels, while promoting regulatory T cell (Treg) differentiation, thereby restoring immune homeostasis[23]. Collectively, these findings suggest that gut microbiota contribute to HT pathogenesis by modulating immune and metabolic pathways, and may also represent promising targets for future therapeutic interventions.
2.4. Gut Microbiota and Thyroid Cancer: Mechanisms and Clinical Evidence
In recent years, increasing attention has been paid to the relationship between gut microbiota and thyroid cancer. Studies indicate that microbial dysbiosis is not only closely associated with thyroid cancer development but also with thyroid nodules and several thyroid function indices. In a cohort of 74 participants, thyroid tumor patients exhibited significantly lower gut microbial diversity compared with healthy controls, with elevated Neisseria and Streptococcus and reduced Butyricimonas and Lactobacillus abundance[24]. These alterations suggest that dysbiosis may disrupt immune homeostasis and local metabolic environments, thereby promoting tumorigenesis. Further evidence links specific microbial compositions to thyroid cancer risk. Metagenomic sequencing revealed enrichment of Prevotella and Fusobacterium in papillary thyroid carcinoma (PTC), while butyrate-producing genera such as Faecalibacterium and Roseburia were depleted, with abundance shifts correlating with tumor markers and tumor size[25]. Microbial metabolites are also implicated: short-chain fatty acids (SCFAs) promote regulatory T cell differentiation and suppress inflammation, whereas lipopolysaccharides (LPS) activate chronic inflammation via the TLR4 pathway, thereby enhancing tumor-related immune responses[26]. In PTC patients, serum propionate and butyrate levels were reduced, while LPS levels increased, correlating positively with disease progression [27]. Microbiota may also modulate immune checkpoint regulation in thyroid cancer. A clinical study reported that Bacteroides abundance was elevated in PD-1/PD-L1–positive patients, while probiotic genera were depleted, suggesting that dysbiosis may shape the tumor immune microenvironment and influence immunotherapy response[28]. Moreover, in patients with concurrent thyroid dysfunction, elevated Bacteroides and reduced Lactobacillus abundance were associated with prolonged disease course and poorer prognosis[29]. Nutritional metabolism provides another link, as gut microbiota can influence iodine absorption and utilization, which are critical for thyroid hormone synthesis. In animal models, high-iodine diets combined with dysbiosis significantly accelerated tumor growth and follicular cell proliferation[30]. Thyroid cancer is also frequently accompanied by metabolic syndrome features such as obesity and insulin resistance. Patients exhibited gut microbiota profiles partially overlapping with those of diabetes, with Ruminococcus abundance correlating with the degree of insulin resistance, suggesting that metabolic disturbances may accelerate carcinogenesis via the gut–thyroid axis[31]. Inflammation represents an additional mechanism. A Chinese cohort study found elevated Escherichia coli abundance in PTC patients, which correlated with serum IL-6, TNF-α levels, and tumor stage, supporting a role of chronic inflammation in tumor aggressiveness[32]. Longitudinal follow-up further showed that patients with recurrence had distinct microbial profiles, with reduced Clostridium cluster XIVa abundance strongly associated with increased recurrence risk[33]. These findings also highlight the diagnostic and classification potential of microbial features. One study proposed a “microbiota risk score” to predict thyroid cancer risk, which outperformed some traditional biochemical markers in differentiating benign nodules from PTC[34]. Intervention studies have shown promise as well: animal experiments demonstrated that probiotic supplementation partially restored microbial homeostasis and reduced thyroid tissue inflammation[35]. Microbiota-targeted strategies, combined with immunotherapy or radioactive iodine therapy, may represent future directions for personalized management. In summary, gut microbiota plays multifaceted roles in the initiation, progression, and prognosis of thyroid cancer, involving immune modulation, metabolic alteration, nutrient absorption, and inflammatory pathways (Figure 3). Identification of specific taxa and metabolites not only deepens mechanistic understanding but also provides new opportunities for risk stratification, diagnosis, and individualized therapeutic intervention.
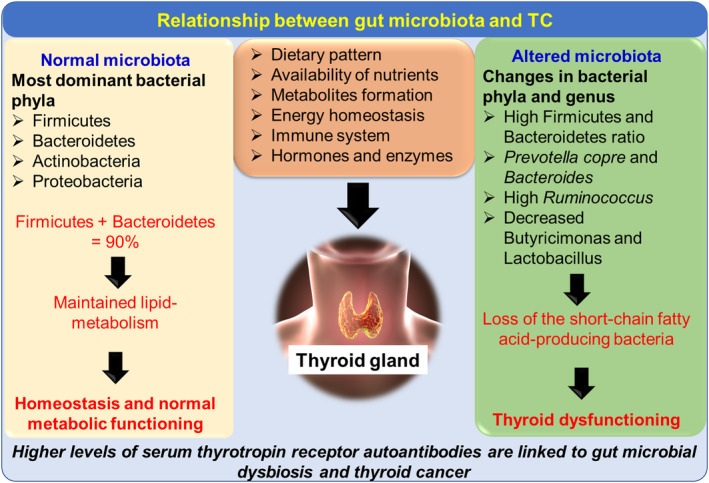
Figure 3. The influence of different gut microbes on thyroid gland function and complications associated with TC[33] (licensed under CC BY 4.0.).
2.5. Gut Microbiota and the Impact on Thyroid Drug Therapy
The role of gut microbiota in thyroid pharmacotherapy is receiving growing attention. Increasing evidence suggests that the intestinal microenvironment not only affects thyroid hormone metabolism and homeostasis but may also directly modulate drug absorption, transport, and efficacy, thereby serving as a critical determinant of personalized treatment. Levothyroxine (LT4) absorption is closely linked to gut microbiota. Clinical studies have shown that reduced abundance of probiotics such as Lactobacillus and Bifidobacterium lowers intestinal LT4 absorption efficiency, whereas enrichment of Bacteroides may indirectly influence drug solubility and bioavailability through alterations in bile acid metabolism[36]. In some patients, such dysbiosis manifests as inadequate response to standard LT4 dosing. Similarly, the metabolism of antithyroid drugs such as methimazole and propylthiouracil is influenced by gut microbes. Certain enzyme-producing strains can alter drug conversion rates and the distribution of metabolites within the intestinal lumen, thereby modulating therapeutic efficacy and adverse effects[37, 38]. Gut microbiota also indirectly affect drug responses by regulating micronutrient absorption. Iodine and selenium are essential for thyroid hormone synthesis and antioxidant defense, and their bioavailability depends on gut microbial balance. Dysbiosis may impair selenium absorption, thereby reducing the metabolic stability of LT4 and weakening its therapeutic effect[39]. Additionally, chronic inflammation driven by abnormal microbial composition may increase LT4 dosage requirements; studies have linked this phenomenon to elevated proportions of Gram-negative bacteria and higher serum LPS levels[40]. Probiotic interventions have shown clinical potential in addressing these challenges. Specific strains such as Lactiplantibacillus plantarum and Bifidobacterium longum have been demonstrated to improve thyroid function, shorten TSH recovery time, and enhance drug efficacy[41]. Meanwhile, fecal microbiota transplantation (FMT) has been explored as an emerging strategy to improve antithyroid drug tolerance and efficacy. Animal experiments indicate that FMT can restore SCFA levels, modulate immune responses, and rebalance gut–thyroid axis signaling pathways, thereby enhancing therapeutic outcomes[42]. Overall, the influence of gut microbiota on thyroid pharmacotherapy is multifaceted, encompassing drug absorption, metabolism, immune regulation, and micronutrient utilization. Future strategies combining probiotics, dietary modulation, or FMT may further improve the efficacy and safety of thyroid medications and advance the development of personalized treatment.
3. Gut Microbiota as Diagnostic and Prognostic Biomarkers in Thyroid Diseases
In recent years, increasing evidence has suggested that gut microbiota and their metabolites are not only associated with the onset of thyroid diseases but may also serve as potential diagnostic and prognostic biomarkers. Their functional roles involve immune regulation, inflammatory pathways, and metabolic networks, with several studies proposing specific microbial signatures as indicators for disease stratification or risk prediction. Thyroid cancer has yielded the most consistent and clinically translatable findings. Elevated abundance of Neisseria and Streptococcus has been strongly linked to tumorigenesis, and their combined pattern effectively discriminates patients with cancer from healthy individuals, suggesting potential value in early diagnosis[43]. In addition, changes in gut microbiota–derived metabolites such as reduced short-chain fatty acids (SCFAs) and elevated lipopolysaccharides (LPS) have been associated with tumor stage and inflammatory status[44], reflecting both disease progression and immune microenvironmental disruption. Other reports have shown that high abundance of Clostridium correlates with lymph node metastasis in papillary thyroid carcinoma (PTC), while loss of Ruminococcus is more frequent in early-stage disease, offering clues for staging and invasion assessment[45]. Long-term follow-up further revealed that patients with persistent high levels of Streptococcus but lacking Lactobacillus after surgery were at significantly higher risk of recurrence[46], highlighting microbiota features as potential predictors of relapse. In autoimmune thyroid diseases (AITD), characteristic microbial signatures have also been observed. In Graves' disease, Prevotella abundance was significantly correlated with thyrotropin receptor antibody (TRAb) levels, suggesting its value as an indicator of disease activity[3]. In HT, reduction of butyrate-producing bacteria was closely associated with elevated serum TSH and increased thyroid parenchymal destruction, suggesting a potential marker of disease severity. In thyroid dysfunction, primary hypothyroidism has been linked to impaired SCFA production and elevated LPS levels, which not only correlate with thyroid hormone deficiency but also serve as sensitive metabolic markers for distinguishing patients from controls[47]. In hyperthyroidism, enrichment of Clostridium was positively correlated with serum FT4 levels and proposed as an auxiliary diagnostic indicator[48]. Microbial features may also predict treatment responses. Studies have shown that patients with poor levothyroxine absorption often present with Lactobacillus depletion and impaired intestinal barrier function, highlighting microbiota features as indicators of reduced drug bioavailability[49]. In Graves' disease, responsiveness to antithyroid drugs has been associated with microbial structure, where the abundance of specific taxa could identify patients at risk of relapse[50]. Moreover, restoration of microbial balance following probiotic supplementation correlated positively with TSH improvement, supporting its role as a potential marker for therapeutic monitoring[51]. Overall, existing studies provide quantifiable diagnostic and prognostic insights based on microbial structure, functional metabolites (SCFAs, LPS), and clinical outcomes (treatment efficacy, recurrence risk). Among these, thyroid cancer–related findings demonstrate the greatest translational potential, with the prospect of microbiota-based multidimensional models enabling early detection, staging evaluation, and recurrence monitoring. Nevertheless, large-scale, prospective studies remain necessary to validate their reliability and generalizability. Future work should focus on integrating microbial biomarkers into clinical decision-making pathways and testing their utility in combination with molecular and imaging approaches, in order to accelerate their translation into precision medicine for thyroid diseases.
4. Gut Microbiota Interventions in the Management of Thyroid Diseases
In recent years, the potential value of gut microbiota–targeted interventions in thyroid disease management, particularly thyroid cancer, has gained increasing attention. Both clinical and experimental studies suggest that modulation of the microbiota not only influences the development of thyroid dysfunction but may also improve therapeutic efficacy and prognosis. For example, probiotic supplementation has been shown to alleviate constipation, fatigue, and metabolic disturbances induced by thyroid hormone withdrawal (THW) after thyroidectomy, while reducing serum and fecal LPS levels, thereby improving intestinal barrier function and immune homeostasis[52]. In Graves' disease, probiotic intervention has been associated with reductions in thyrotropin receptor antibody (TRAb) levels, indicating an immunomodulatory potential that may help lower relapse risk[53]. Diet represents another important means of microbiota modulation. High-fiber diets promote the production of short-chain fatty acids (SCFAs), improve inflammatory status, and may enhance the metabolic stability of thyroid hormones[54]. In contrast, high-fat diets have been linked to expansion of pro-inflammatory taxa and increased risk of thyroid tumor progression[55]. Micronutrients such as selenium and zinc also influence microbial diversity and antioxidant pathways, potentially improving immune responses and treatment tolerance in patients with thyroid dysfunction[56]. Although direct evidence for fecal microbiota transplantation (FMT) in thyroid disease is limited, its demonstrated ability to enhance responses to immune checkpoint inhibitors in other cancers[57] provides a rationale for exploring its application in thyroid cancer, particularly for overcoming resistance and improving prognosis. Prognostic associations between microbiota and clinical outcomes are increasingly recognized. Gut microbial composition has been correlated with responses to radioactive iodine therapy[58]; depletion of Lactobacillus has been linked to poor levothyroxine absorption and drug intolerance, serving as a predictor of therapeutic outcomes[59]; reduced microbial diversity following surgery or radiotherapy has been associated with increased recurrence risk[60]; and probiotic supplementation has been reported to restore microbial balance and coincide with TSH recovery, supporting its role as a potential monitoring tool[61]. Furthermore, FMT has already demonstrated efficacy in improving checkpoint inhibitor responses in melanoma[62], suggesting translational potential for thyroid cancer. Overall, evidence from microbial composition, functional metabolites (SCFAs, LPS), and clinical endpoints (treatment response, recurrence risk) highlights the promise of microbiota-based interventions. These findings suggest that targeted modulation of gut microbiota holds significant potential for precision stratification and personalized management of thyroid cancer, though large-scale prospective studies are needed to validate their clinical utility. Future efforts should aim to integrate microbiota-based biomarkers into standardized clinical workflows and explore synergistic strategies combining microbiota modulation with conventional and novel therapies, in order to translate mechanistic insights into tangible benefits for patients.
5. Controversies and Challenges: Causality, Heterogeneity, and Insufficient Evidence
Despite increasing links between gut microbiota and thyroid diseases, the translational pathway from correlation to clinical application still faces three major challenges: unclear causality, substantial heterogeneity, and methodological limitations with insufficient levels of evidence. These issues are not unique to thyroid cancer (TC) but also apply to autoimmune thyroid diseases (AITD, including Graves' disease and Hashimoto's thyroiditis) and thyroid dysfunction (hyper- and hypothyroidism).
5.1. Unclear causality and unaddressed confounding
Most studies are cross-sectional or rely on postoperative samples, making it difficult to distinguish whether dysbiosis is a causal factor or a secondary effect of disease and treatment[63]. Hyperthyroidism accelerates intestinal transit and hypothyroidism delays motility, both independently altering fecal microbiota and complicating the attribution to “primary microbial effects”[64]. Perioperative factors, radioactive iodine (RAI), levothyroxine replacement, antithyroid drugs, proton pump inhibitors, and antibiotic use can all disrupt the microbiota through bile acid and mucosal immune pathways[65, 66]. Without sensitivity analyses or instrumental variable approaches to account for drug exposures, dietary intake (iodine, selenium, fiber), and lifestyle factors, statistical models are prone to overestimating microbiota–outcome associations[67]. Prospective inception cohorts, time-series sampling, and causal inference approaches (including Mendelian randomization and intervention–response studies) are needed to clarify directionality[68].
5.2. Dual heterogeneity across populations and disease subtypes
Within thyroid cancer, molecular pathways, immune microenvironments, and treatment responses vary considerably across papillary, follicular, medullary, and anaplastic subtypes; pooled analyses risk signal dilution and poor reproducibility[69]. AITD also shows heterogeneity in antibody profiles (TRAb, TPOAb, TgAb) and disease activity, while metabolic states, body composition, and comorbidities (obesity, type 2 diabetes) can systematically shift microbiota in hyper- and hypothyroid patients[70]. Cross-regional and ethnic differences in diet and lifestyle (e.g., iodine/selenium intake, fermented foods, cereal fiber) further reduce biomarker stability across external cohorts[71]. Subtype-specific modeling and cross-center validation are therefore prerequisites for clinical utility[72].
5.3. Technical and methodological biases
Low-biomass samples (thyroid tissue, blood) are highly vulnerable to environmental or reagent contamination; without negative controls, decontamination, and absolute quantification (e.g., exogenous standards or qPCR), false-positive microbial signals may occur[73]. Differences in resolution and functional inference between 16S rRNA sequencing and metagenomics, as well as variability in DNA extraction protocols, sequencing platforms, batch effects, and bioinformatics pipelines (OTU/ASV construction, host-read removal), can introduce systematic error[74]. Many models with apparently high AUCs rely only on internal cross-validation and lack preregistration or multicenter replication, raising concerns about clinical generalizability[75].
5.4. Incomplete mechanistic chains
Most evidence in TC and AITD remains “structural associations” without integrated demonstration across the full continuum: microbiota → metabolites (SCFAs, secondary bile acids, indoles) → host pathways (deiodinase activity, FXR/PXR, Treg/Th17 balance) → clinical phenotypes, within the same cohort[76]. Reversibility and dose–response data are also limited; whether targeted supplementation of butyrate producers or Akkermansia can consistently increase metabolite levels and improve imaging or pathological endpoints remains to be tested in randomized controlled trials[77]. In AITD, stratified assessment of “microbiota modulation–antibody titers–relapse/remission” over time is required; in thyroid dysfunction, distinguishing the marginal contribution of “hormone correction” versus “microbiota intervention” is essential[63, 68].
5.5. Clinical benefit and safety margins remain uncertain
On diagnosis, some fecal microbial features improve discrimination of benign versus malignant nodules or PTC staging, but the incremental benefit over fine-needle aspiration cytology (FNAC) and molecular assays (BRAF/TERT) has not been systematically quantified; most models lack joint evaluations with imaging or molecular tools[78]. On prognosis, recurrence models that omit stage, RAI dose, TSH suppression intensity, and molecular subtype may exaggerate the “independent effect” of microbiota[64, 74]. On interventions, although probiotics, diet, and FMT have shown efficacy in improving immunotherapy responses across other cancers, these findings cannot be directly extrapolated to TC, where surgery and RAI remain the mainstay. Moreover, risks of FMT—including infection, transfer of antibiotic resistance genes, and immune-related adverse events—must be carefully considered in vulnerable groups such as AITD or thyroid storm patients[79].
5.6. Toward clinical translation
Future research should advance along three parallel lines: (i) methodology—developing standardized SOPs (sampling, storage, sequencing, analysis), implementing negative controls and absolute quantification, multicenter preregistration, and decision curve analyses to report true clinical gain[67, 71]; (ii) causality and mechanisms—prospective inception cohorts with multi-omics (metagenomics, metabolomics, immunomics, serum iodine/selenium), combined with MR and small-scale “mechanistic RCTs” targeting butyrate producers or Akkermansia to test reversibility and dose–response[80]; (iii) clinical embedding—nested designs in AITD remission/relapse, hyper- and hypothyroidism dose adjustments, and perioperative/RAI contexts in thyroid cancer, using translatable endpoints (recurrence-free survival, RAI sensitivity, antibody titers/TSH dynamics), integrated with cost–benefit analyses to confirm real-world clinical value[81].
In summary, these challenges do not negate the role of gut microbiota in thyroid diseases but emphasize that its integration into clinical practice requires filling causal gaps, addressing heterogeneity and methodological biases, and demonstrating incremental value beyond current standards across real-world endpoints. This consensus applies broadly to thyroid cancer, AITD, and thyroid dysfunction.
6. Conclusion
With the expanding depth of gut microbiota research, evidence increasingly indicates that microbial communities are critically involved in the onset, progression, and management of thyroid diseases. Beyond their associations with thyroid hormone metabolism, autoimmune responses, and inflammatory pathways, particular attention is now turning to their potential roles in thyroid cancer initiation and progression. Microbiota-derived signatures and metabolites may provide not only diagnostic and prognostic biomarkers but also mechanistic insight into tumor development and therapeutic responsiveness. Nevertheless, most current findings are derived from cross-sectional studies, limiting causal inference and leaving clinical translation in its infancy. Future priorities include establishing longitudinal, multicenter cohorts to clarify how microbial profiles shape the natural history of thyroid cancer and autoimmune thyroid disorders; integrating multi-omics with functional validation to delineate microbial metabolite–tumor–immune interactions; and advancing interventional trials in real-world settings to test strategies such as probiotics, dietary modulation, and fecal microbiota transplantation (FMT). Persistent challenges—uncertain causal chains, inconsistent findings across populations, and insufficient mechanistic evidence—remain, but these gaps define clear opportunities for progress. Ultimately, bridging mechanistic research and clinical application in a reproducible, translatable manner will determine whether microbiota-based biomarkers and interventions can be fully incorporated into precision medicine for thyroid cancer and related thyroid diseases.
Conflicts of Interest
The authors declare no conflict of interest.
Funding
No funding.
Author Contributions
All authors read and approved the final manuscript.
Acknowledgements
This study is grateful to all the authors for their full contributions.
References
-
L. Fang, J. Ning. “Recent advances in gut microbiota and thyroid disease: Pathogenesis and therapeutics in autoimmune, neoplastic, and nodular conditions.” Front. Cell. Infect. Microbiol. 2024, 14, 1465928.
-
M. E. Ludgate, G. Masetti, P. Soares. “The relationship between the gut microbiota and thyroid disorders.” Nat. Rev. Endocrinol. 2024, 20, 9, 511–25.
-
Y. Fang, X. Zhang, R. Huang, J. Liu, Z. Li. “Gut microbiota and autoimmune thyroid disease: A bidirectional Mendelian randomization study and mediation analysis.” Front. Microbiol. 2024, 15, 1443643.
-
L. Sessa, E. Malavolta, G. Sodero, C. Cipolla, D. Rigante. “The conspiring role of gut microbiota as primer of autoimmune thyroid diseases: A scoping focus.” Autoimmun. Rev. 2025, 24, 5, 103780.
-
K. Yan, X. Sun, C. Fan, X. Wang, H. Yu. “Unveiling the role of gut microbiota and metabolites in autoimmune thyroid diseases: Emerging perspectives.” Int. J. Mol. Sci. 2024, 25, 20, 10918.
-
I. Legakis, G. P. Chrousos, S. Chatzipanagiotou. “Thyroid diseases and intestinal microbiome.” Horm. Metab. Res. 2023, 55, 12, 813–818.
-
S. L. Collins, J. G. Stine, J. E. Bisanz, C. D. Okafor, A. D. Patterson. “Bile acids and the gut microbiota: Metabolic interactions and impacts on disease.” Nat. Rev. Microbiol. 2023, 21, 4, 236–247.
-
C. Shi, J. Chen, S. He, Y. Zhang, Y. Zhang, L. Yu. “Cross-talk between the gut microbiota and hypothyroidism: A bidirectional two-sample Mendelian randomization study.” Front. Nutr. 2024, 11, 1286593.
-
J. Knezevic, C. Starchl, A. Tmava Berisha, K. Amrein. “Thyroid-gut-axis: How does the microbiota influence thyroid function?” Nutrients 2020, 12, 6, 1769.
-
S. C. Russo, F. Salas-Lucia, A. C. Bianco. “Deiodinases and the metabolic code for thyroid hormone action.” Endocrinology 2021, 162, 8, bqab059.
-
W. Jiang, X. Yu, R. O. Kosik, Y. Song, T. Qiao, J. Tong. “Gut microbiota may play a significant role in the pathogenesis of Graves’ disease.” Thyroid 2021, 31, 5, 810–820.
-
M. Yang, B. Sun, J. Li, B. Yang, J. Xu, X. Zhou, J. Yu, X. Zhang, Q. Zhang, S. Zhou, X. Sun. “Alteration of the intestinal flora may participate in the development of Graves’ disease: A study conducted among the Han population in southwest China.” Endocr. Connect. 2019, 8, 7, 822–828.
-
Q. Zhu, Q. Hou, S. Huang, Q. Ou, D. Huo, Y. Vázquez-Baeza, C. Cen, V. Cantu, M. Estaki, H. Chang, P. Belda-Ferre, H.-C. Kim, K. Chen, R. Knight, J. Zhang. “Compositional and genetic alterations in Graves’ disease gut microbiome reveal specific diagnostic biomarkers.” ISME J. 2021, 15, 11, 3399–3411.
-
J. Cao, N. Wang, Y. Luo, C. Ma, Z. Chen, C. Chenzhao, F. Zhang, X. Qi, W. Xiong. “A cause–effect relationship between Graves’ disease and the gut microbiome contributes to the thyroid–gut axis: A bidirectional two-sample Mendelian randomization study.” Front. Immunol. 2023, 14, 977587.
-
H. Chen, J. Cao, F. Zhang, W. Xiong. “Significance of gut microbiota on Graves’ disease.” Int. J. Gen. Med. 2024, 17, 3967–3974.
-
H. Liu, H. Liu, C. Liu, M. Shang, T. Wei, P. Yin. “Gut microbiome and the role of metabolites in the study of Graves’ disease.” Front. Mol. Biosci. 2022, 9, 841223.
-
X. Zhang, Q. Pan, G. Yao, D. Kong, H. Chen, Q. Zhang, Z. Wang. “Bacteroides fragilis and propionate synergize with low-dose methimazole to treat Graves’ disease.” Microbiol. Spectr. 2025, 13, 6, e03186-24.
-
F. Zhao, J. Feng, J. Li, L. Zhao, Y. Liu, H. Chen, Y. Jin, B. Q. Zhu, Y. W. Wei. “Alterations of the gut microbiota in Hashimoto’s thyroiditis patients.” Thyroid 2018, 28, 2, 175–186.
-
M. N. Quraishi, M. Yalchin, C. Blackwell, J. Segal, N. Sharma, P. Hawkey, V. McCune, A. L. Hart, D. Gaya, N. J. Ives, L. Magill, S. Loi, C. Hewitt, K. Gerasimidis, N. J. Loman, R. Hansen, C. McMullan, J. Mathers, C. Quince, N. Crees, T. Iqbal. “STOP-Colitis pilot trial protocol: A prospective, open-label, randomised pilot study to assess two possible routes of faecal microbiota transplant delivery in patients with ulcerative colitis.” BMJ Open 2019, 9, 11, e030659.
-
M. Li, K. Chen, Y. Chen, L. Zhang, Y. Cui, F. Xiao, Z. Liu, W. Zhang, J. Jiang, Q. Zhou, J. Yan, Y. Sun, F. Guan. “Integrative analysis of gut microbiome and host transcriptome reveal novel molecular signatures in Hashimoto’s thyroiditis.” J. Transl. Med. 2024, 22, 1, 1045.
-
C. Virili, P. Fallahi, A. Antonelli, S. Benvenga, M. Centanni. “Gut microbiota and Hashimoto's thyroiditis.” Rev. Endocr. Metab. Disord. 2018, 19, 4, 293–300.
-
K. Mori, Y. Nakagawa, H. Ozaki. “Does the gut microbiota trigger Hashimoto's thyroiditis?” Discov. Med. 2012, 14, 78, 321–326.
-
L. C. F. Cayres, L. V. V. de Salis, G. S. P. Rodrigues, A. v. H. Lengert, A. P. C. Biondi, L. D. B. Sargentini, J. L. Brisotti, E. Gomes, G. L. V. de Oliveira. “Detection of alterations in the gut microbiota and intestinal permeability in patients with Hashimoto thyroiditis.” Front. Immunol. 2021, 12, 579140.
-
X. Yu, W. Jiang, R. O. Kosik, Y. Song, Q. Luo, T. Qiao, J. Tong, S. Liu, C. Deng, S. Qin, Z. Lv, D. Li. “Gut microbiota changes and its potential relations with thyroid carcinoma.” J. Adv. Res. 2022, 35, 61–70.
-
Q. Liu, W. Sun, H. Zhang. “Interaction of gut microbiota with endocrine homeostasis and thyroid cancer.” Cancers (Basel) 2022, 14, 11, 2656.
-
T. Hou, Q. Wang, H. Dai, Y. Hou, J. Zheng, T. Wang, H. Lin, S. Wang, M. Li, Z. Zhao, Y. Chen, Y. Xu, J. Lu, R. Liu, G. Ning, W. Wang, M. Xu, Y. Bi. “Interactive association between gut microbiota and thyroid cancer.” Endocrinology 2023, 165, 1, bqad184.
-
X. Sun, S. Chen, S. Zhao, J. Wang, H. Cheng. “Causal relationship of genetically predicted gut microbiota with thyroid cancer: A bidirectional two-sample Mendelian randomization study.” Front. Endocrinol. (Lausanne) 2024, 15, 1284472.
-
Y. Ren, Y. Zhang, Y. Cheng, H. Qin, H. Zhao. “Genetic liability of gut microbiota for idiopathic pulmonary fibrosis and lung function: A two-sample Mendelian randomization study.” Front. Cell Infect. Microbiol. 2024, 14, 1348685.
-
K.-H. Kim, S. J. Lee, J. Kim, Y. Moon. “Leveraging xenobiotic-responsive cancer stemness in cell line-based tumoroids for evaluating chemoresistance: A proof-of-concept study on environmental susceptibility.” Int. J. Mol. Sci. 2024, 25, 21, 11383.
-
Y. Wang, M. Bai, Q. Peng, L. Li, F. Tian, Y. Guo, C. Jing. “Angiogenesis, a key point in the association of gut microbiota and its metabolites with disease.” Eur. J. Med. Res. 2024, 29, 1, 614.
-
J. Zhou, X. Zhang, Z. Xie, Z. Li. “Exploring reciprocal causation: Bidirectional Mendelian randomization study of gut microbiota composition and thyroid cancer.” J. Cancer Res. Clin. Oncol. 2024, 150, 2, 75.
-
Z. Quan, X. Zhang, S. Wang, Y. Meng. “Causal analysis of the gut microbiota in differentiated thyroid carcinoma: a two-sample Mendelian randomization study.” Front. Genet. 2023, 14, 1299930.
-
Y. Kun, X. Wei, H. Wang, X. Nie, Q. Dai. “Exploring the oral-gut microbiota during thyroid cancer: Factors affecting the thyroid functions and cancer development.” Food Sci. Nutr. 2023, 11, 10, 5657–5674.
-
J. Zhang, F. Zhang, C. Zhao, Q. Xu, C. Liang, Y. Yang, H. Wang, Y. Shang, Y. Wang, X. Mu, D. Zhu, C. Zhang, J. Yang, M. Yao, L. Zhang. “Dysbiosis of the gut microbiome is associated with thyroid cancer and thyroid nodules and correlated with clinical index of thyroid function.” Endocrine 2019, 64, 3, 564–574.
-
J. Jiao, Y. Zheng, Q. Zhang, D. Xia, L. Zhang, N. Ma. “Saliva microbiome changes in thyroid cancer and thyroid nodules patients.” Front. Cell Infect. Microbiol. 2022, 12, 989188.
-
Y. Xie, F. Hu, D. Xiang, H. Lu, W. Li, A. Zhao, L. Huang, R. Wang. “The metabolic effect of gut microbiota on drugs.” Drug Metab. Rev. 2020, 52, 1, 139–156.
-
Z. Yao, M. Zhao, Y. Gong, W. Chen, Q. Wang, Y. Fu, T. Guo, J. Zhao, L. Gao, T. Bo. “Relation of gut microbes and L-thyroxine through altered thyroxine metabolism in subclinical hypothyroidism subjects.” Front. Cell Infect. Microbiol. 2020, 10, 495.
-
J. Zhang, J. Zhang, R. Wang. “Gut microbiota modulates drug pharmacokinetics.” Drug Metab. Rev. 2018, 50, 3, 357–368.
-
C. Virili, N. Brusca, S. Capriello, M. Centanni. “Levothyroxine therapy in gastric malabsorptive disorders.” Front. Endocrinol. (Lausanne) 2021, 11, 621616.
-
J. Sun, F. Zhao, B. Lin, J. Feng, X. Wu, Y. Liu, L. Zhao, B. Zhu, Y. Wei. “Gut microbiota participates in antithyroid drug induced liver injury through the lipopolysaccharide related signaling pathway.” Front. Pharmacol. 2020, 11, 598170.
-
M. Wu, C. Chi, Y. Yang, S. Guo, T. Li, M. Gu, T. Zhang, H. Gao, R. Liu, C. Yin. “Dynamics of gut microbiota during pregnancy in women with TPOAb-positive subclinical hypothyroidism: a prospective cohort study.” BMC Pregnancy Childbirth 2022, 22, 1, 592.
-
X. Su, X. Yin, Y. Liu, X. Yan, S. Zhang, X. Wang, Z. Lin, X. Zhou, J. Gao, Z. Wang, Q. Zhang. “Gut dysbiosis contributes to the imbalance of Treg and Th17 cells in Graves' disease patients by propionic acid.” J. Clin. Endocrinol. Metab. 2020, 105, 11, dgaa511.
-
B. Gong, C. Wang, F. Meng, H. Wang, B. Song, Y. Yang, Z. Shan. “Association between gut microbiota and autoimmune thyroid disease: a systematic review and meta-analysis.” Front. Endocrinol. (Lausanne) 2021, 12, 774362.
-
J. K. Paul, M. Azmal, A. S. N. Been Haque, M. Meem, O. F. Talukder, A. Ghosh. “Unlocking the secrets of the human gut microbiota: comprehensive review on its role in different diseases.” World J. Gastroenterol. 2025, 31, 5, 99913.
-
K. Yan, X. Sun, C. Fan, X. Wang, H. Yu. “Unveiling the role of gut microbiota and metabolites in autoimmune thyroid diseases: emerging perspectives.” Int. J. Mol. Sci. 2024, 25, 20, 10918.
-
P. Bargiel, M. Szczuko, L. Stachowska, P. Prowans, N. Czapla, M. Markowska, J. Petriczko, J. Kledzik, A. Jędrzejczyk-Kledzik, J. Palma, P. Zabielska, D. Maciejewska-Markiewicz. “Microbiome metabolites and thyroid dysfunction.” J. Clin. Med. 2021, 10, 16, 3609.
-
J. Zhou, X. Zhang, Z. Xie, Z. Li. “Exploring reciprocal causation: bidirectional Mendelian randomization study of gut microbiota composition and thyroid cancer.” J. Cancer Res. Clin. Oncol. 2024, 150, 2, 75.
-
H.-S. Ejtahed, P. Angoorani, A.-R. Soroush, S.-D. Siadat, N. Shirzad, S. Hasani-Ranjbar, B. Larijani. “Our little friends with big roles: Alterations of the gut microbiota in thyroid disorders.” Endocr. Metab. Immune Disord. Drug Targets 2020, 20, 3, 344–350.
-
X. Zhu, C. Zhang, S. Feng, R. He, S. Zhang. “Intestinal microbiota regulates the gut-thyroid axis: the new dawn of improving Hashimoto thyroiditis.” Clin. Exp. Med. 2024, 24, 1, 39.
-
F. Biscarini, G. Masetti, I. Muller, H. L. Verhasselt, D. Covelli, G. Colucci, L. Zhang, M. S. Draman, O. Okosieme, P. Taylor, C. Daumerie, M. C. Burlacu, M. Marinò, D. G. Ezra, P. Perros, S. Plummer, A. Eckstein, M. Salvi, J. R. Marchesi, M. Ludgate. “Gut microbiome associated with Graves disease and Graves orbitopathy: The INDIGO multicenter European study.” J. Clin. Endocrinol. Metab. 2023, 108, 8, 2065–2077.
-
D. Huo, C. Cen, H. Chang, Q. Ou, S. Jiang, Y. Pan, K. Chen, J. Zhang. “Probiotic Bifidobacterium longum supplied with methimazole improved the thyroid function of Graves' disease patients through the gut-thyroid axis.” Commun. Biol. 2021, 4, 1, 1046.
-
B. Lin, F. Zhao, Y. Liu, X. Wu, J. Feng, X. Jin, W. Yan, X. Guo, S. Shi, Z. Li, L. Liu, H. Chen, H. Wang, S. Wang, Y. Lu, Y. Wei. “Randomized clinical trial: Probiotics alleviated oral-gut microbiota dysbiosis and thyroid hormone withdrawal-related complications in thyroid cancer patients before radioiodine therapy following thyroidectomy.” Front. Endocrinol. (Lausanne) 2022, 13, 834674.
-
Q. Shu, C. Kang, J. Li, Z. Hou, M. Xiong, X. Wang, H. Peng. “Effect of probiotics or prebiotics on thyroid function: A meta-analysis of eight randomized controlled trials.” PLoS One 2024, 19, 1, e0296733.
-
M. Wang, Y. Zhu. “Gut microbiome versus thyroid cancer: Association and clinical implications (Review).” Oncol. Lett. 2025, 30, 1, 368.
-
L.-Y. Zhao, J.-X. Mei, G. Yu, L. Lei, W.-H. Zhang, K. Liu, X.-L. Chen, D. Kołat, K. Yang, J.-K. Hu. “Role of the gut microbiota in anticancer therapy: from molecular mechanisms to clinical applications.” Signal Transduct. Target Ther. 2023, 8, 201.
-
A. C. Fenneman, E. Bruinstroop, M. Nieuwdorp, A. H. van der Spek, A. Boelen. “A comprehensive review of thyroid hormone metabolism in the gut and its clinical implications.” Thyroid 2023, 33, 1, 32–44.
-
E. N. Baruch, I. Youngster, G. Ben-Betzalel, R. Ortenberg, A. Lahat, L. Katz, K. Adler, D. Dick-Necula, S. Raskin, N. Bloch, D. Rotin, L. Anafi, C. Avivi, J. Melnichenko, Y. Steinberg-Silman, R. Mamtani, H. Harati, N. Asher, R. Shapira-Frommer, T. Brosh-Nissimov, Y. Eshet, S. Ben-Simon, O. Ziv, M. A. W. Khan, M. Amit, N. J. Ajami, I. Barshack, J. Schachter, J. A. Wargo, O. Koren, G. Markel, B. Boursi. “Fecal microbiota transplant promotes response in immunotherapy-refractory melanoma patients.” Science 2021, 371, 6529, 602–609.
-
N. Gronich, I. Lavi, G. Rennert, W. Saliba. “Cancer risk after radioactive iodine treatment for hyperthyroidism: A cohort study.” Thyroid 2020, 30, 2, 243–250.
-
C. H. Woelk, A. Snyder. “Modulating gut microbiota to treat cancer.” Science 2021, 371, 6529, 573–574.
-
A. Abubaker, Z. Neale, Y. Hoballah, Patient Mosaic Team, Facilitating Anaplastic Thyroid Cancer Specialized Treatment Consortium, J. Wargo, S. Lai, N. Ajami, A. Maniakas. “Clinical implications of the gut microbiome in anaplastic thyroid cancer.” Endocrine Abstracts 2024, 101, OP06-06.
-
M. Karimi, R. Rabiei, K. Kazemi, R. Bagheri Motlagh, O. Asbaghi. “Effects of probiotics and synbiotics oral supplementation on thyroid function in adults: a grade-assessed systematic review and meta-analysis.” Thyroid Res. 2025, 18, 39.
-
L. Yuan, P. Yang, G. Wei, X. Hu, S. Chen, J. Lu, L. Yang, X. He, G. Bao. “Tumor microbiome diversity influences papillary thyroid cancer invasion.” Commun. Biol. 2022, 5, 864.
-
I. Vujkovic-Cvijin, J. Sklar, L. Jiang, L. Natarajan, R. Knight, Y. Belkaid. “Host variables confound gut microbiota studies of human disease.” Nature 2020, 587, 7834, 448–454.
-
R. Daher, T. Yazbeck, J. Bou Jaoude, B. Abboud. “Consequences of dysthyroidism on the digestive tract and viscera.” World J. Gastroenterol. 2009, 15, 23, 2834–2838.
-
L. Zheng, L. Zhang, L. Tang, D. Huang, D. Pan, W. Guo, S. He, Y. Huang, Y. Chen, X. Xiao, B. Tang, J. Chen. “Gut microbiota is associated with response to 131I therapy in patients with papillary thyroid carcinoma.” Eur. J. Nucl. Med. Mol. Imaging 2023, 50, 5, 1453–1465.
-
B. V. Jones, M. Begley, C. Hill, C. G. M. Gahan, J. R. Marchesi. “Functional and comparative metagenomic analysis of bile salt hydrolase activity in the human gut microbiome.” Proc. Natl. Acad. Sci. U.S.A. 2008, 105, 36, 13580–13585.
-
I. Vujkovic-Cvijin, J. Sklar, L. Jiang, L. Natarajan, R. Knight, Y. Belkaid. “Host variables confound gut microbiota studies of human disease.” Nature 2020, 587, 7834, 448–454.
-
X. Zhou, X. Shen, J. S. Johnson, D. J. Spakowicz, M. Agnello, W. Zhou, M. Avina, A. Honkala, F. Chleilat, S. J. Chen, K. Cha, S. Leopold, C. Zhu, L. Chen, L. Lyu, D. Hornburg, S. Wu, X. Zhang, C. Jiang, L. Jiang, R. Jian, A. W. Brooks, M. Wang, K. Contrepois, P. Gao, S. M. Schüssler-Fiorenza Rose, T. D. B. Tran, H. Nguyen, A. Celli, B. Y. Hong, E. J. Bautista, Y. Dorsett, P. B. Kavathas, Y. Zhou, E. Sodergren, G. M. Weinstock, M. P. Snyder. “Longitudinal profiling of the microbiome at four body sites reveals core stability and individualized dynamics during health and disease.” Cell Host Microbe 2024, 32, 4, 506–526.e9.
-
J. Hu, I. J. Yuan, S. Mirshahidi, A. Simental, S. C. Lee, X. Yuan. “Thyroid carcinoma: phenotypic features, underlying biology and potential relevance for targeting therapy.” Int. J. Mol. Sci. 2021, 22, 4, 1950.
-
M. Byndloss, S. Devkota, F. Duca, J. H. Niess, M. Nieuwdorp, M. Orho-Melander, Y. Sanz, V. Tremaroli, L. Zhao. “The gut microbiota and diabetes: research, translation, and clinical applications—2023 Diabetes, Diabetes Care, and Diabetologia expert forum.” Diabetes Care 2024, 47, 9, 1491–1508.
-
Q. Y. Ang, D. L. Alba, V. Upadhyay, J. E. Bisanz, J. Cai, H. L. Lee, E. Barajas, G. Wei, C. Noecker, A. D. Patterson, S. K. Koliwad, P. J. Turnbaugh. “The East Asian gut microbiome is distinct from colocalized White subjects and connected to metabolic health.” eLife 2021, 10, e70349.
-
R. Sinha, G. Abu-Ali, E. Vogtmann, A. A. Fodor, B. Ren, A. Amir, E. Schwager, J. Crabtree, S. Ma, The Microbiome Quality Control Project Consortium, C. C. Abnet, R. Knight, O. White, C. Huttenhower. “Assessment of variation in microbial community amplicon sequencing by the Microbiome Quality Control (MBQC) project consortium.” Nat. Biotechnol. 2017, 35, 11, 1077–1086.
-
S. J. Salter, M. J. Cox, E. M. Turek, S. T. Calus, W. O. Cookson, M. F. Moffatt, P. Turner, J. Parkhill, N. J. Loman, A. W. Walker. “Reagent and laboratory contamination can critically impact sequence-based microbiome analyses.” BMC Biol. 2014, 12, 87.
-
M. J. Saadh, O. Q. B. Allela, S. Ballal, M. S. Mahdi, R. Chahar, R. Verma, R. K. A. Al-Hussein, M. Adil, M. J. Jawad, A. M. A. Al-Nuaimi. “The effects of microbiota-derived short-chain fatty acids on T lymphocytes: From autoimmune diseases to cancer.” Semin. Oncol. 2025, 52, 5, 152398.
-
O. Shete, T. S. Ghosh. “Normal gut microbiomes in diverse populations: Clinical implications.” Annu. Rev. Med. 2025, 76, 1, 95–114.
-
B. Ge, R. C. McDonald, Q. Yang, K. J. Domesle, S. Sarria, X. Li, C.-H. Hsu, K. G. Jarvis, D. A. Tadesse. “Exploring animal food microbiomes and resistomes via 16S rRNA gene amplicon sequencing and shotgun metagenomics.” Appl. Environ. Microbiol. 2025, 91, 2, e0223024.
-
C. Depommier, A. Everard, C. Druart, H. Plovier, M. Van Hul, S. Vieira-Silva, G. Falony, J. Raes, D. Maiter, N. M. Delzenne, M. de Barsy, A. Loumaye, M. P. Hermans, J.-P. Thissen, W. M. de Vos, P. D. Cani. “Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: a proof-of-concept exploratory study.” Nat. Med. 2019, 25, 7, 1096–1103.
-
R. C. Simpson, E. Shanahan, R. A. Scolyer, G. V. Long. “Targeting the microbiome to overcome resistance.” Cancer Cell 2021, 39, 2, 151–153.
-
J. Fu, X. Yin, X. Wang, S. Xiao, X. Wu, C. Duan, W. Yu, G. Zhang. “Diagnostic value of FNAC combined with BRAFV600E mutation detection in Hashimoto's thyroiditis complicated with papillary thyroid carcinoma.” Front. Endocrinol. (Lausanne) 2024, 15, 1366724.
-
C. Shi, J. Chen, S. He, Y. Zhang, Y. Zhang, L. Yu. “Cross-talk between the gut microbiota and hypothyroidism: a bidirectional two-sample Mendelian randomization study.” Front. Nutr. 2024, 11, 1286593.